El origen de los eucariontes es un proceso que tiene un origen procariota. Este proceso se produjo por endosimbiosis entre varios organismos procariotas, en donde el ancestro principal protoeucariota es de tipo arqueano y las mitocondrias y cloroplastos son de origen bacteriano. Es discutible la incorporación de otros organismos procariotas. Las procariotas se diferenciaron de las eucariotas porque las procariotas no tienen núcleo y tienen genoma y las eucariotas si. Ejemplo de esto, es que las cianos bacterias entraron en procariotas grandes y la convirtieron en eucariotas, como las algas dieron origen a las plantas.
La tinción de Gram o coloración de Gram es un tipo de tinción diferencial empleado en Bacteriología para la visualización de bacterias, sobre todo en es una prueba potente y rápida que nos permite diferenciar dos clases de bacterias.
Es una prueba potente y rápida que nos permite diferenciar dos clases de bacterias estas son: Bacterias gram positivas y las gram negativas.
Las grampositivas se tiñen de morado ya que el colorante se queda atrapad en la capa gruesa de peptidoglucanos que rodea a la célula
Las gramnegativas tienen una capa de petidoglucano mucho más delgada es por ello que no retiene el violeta cristal y por esto las células se tiñen con safranina y las observamos rojas.
*Bacilos (en forma de bastón, alargados),que a su vez pueden tener varios aspectos:cilíndricos,fusiformes y en forma de maza, etc.
Atendiendo a los tipos de extremos, éstos pueden ser:
*Redondeados (lo más frecuente)
*Cuadrados
*Biselados
*Afilados
*Espirilos: al igual que los bacilos, tienen un eje más largo que otro, pero dicho eje no es recto, sino que sigue una forma de espiral, con una o más de una vuelta de hélice.
*Vibrios: proyectada su imagen sobre el plano tienen forma de coma, pero en el espacio suelen corresponder a una forma espiral con menos de una vuelta de hélice.
Otros tipos de formas son filamentos, ramificados o no,anillos casi cerrados, y formas con prolongaciones (con prostecas).
AGRUPACIONES BACTERIANAS
Las bacterias normalmente se multiplican por fisión transversal binaria. En muchas especies, las células hijas resultantes de un evento de división por fisión tienden a dispersarse por separado al medio, debido a la actuación de fuerzas físicas (movimiento browniano, cizallamiento, corrientes de convección, etc). Esto hace que al observar al microscopio una población de estas bacterias veamos mayoritariamente células aisladas. Pero en algunas especies las células hijas pueden permanecer unidas entre sí (al menos durante un cierto tiempo tras la división de la que proceden) debido a que el tabique sea incompleto o a la existencia de capas mucosas que retienen juntos los productos de la división.
Si la tendencia a permanecer unidas es baja, tendremos agrupaciones de dos células, que dependiendo que sean de morfología esférica o alargada, se denominan como: *Diplococos *Diplobacilos
Si la tendencia a permanecer unidas es mayor (por más tiempo), nos encontramos con varias posibilidades, dependiendo del número de planos de división y de la relación entre ellos:
Si los tabiques son paralelos entre sí (o sea, existe un solo plano de división): estreptococos (cadenetas arrosariadas de cocos) y estreptobacilos (cadenetas de bacilos).
Si existe más de un plano de división, en el caso de cocos podemos encontrar tres posibilidades:
Dos planos perpendiculares: tétradas (4 céls. en un plano) o múltiplos.
Tres planos ortogonales: sarcinas (paquetes cúbicos).
Muchos planos de división: estafilococos (racimos irregulares). En el caso de bacilos, se pueden dar variantes adicionales, debido a la posibilidad de que se produzcan movimientos postfisionales (en algunos casos con desgarro):
Bacilos en empalizada o en paquetes de cigarrillos (debido a giros de 180grado)
Algunos géneros bacterianos se agrupan de una manera característica. Esta agrupación se debe a la tendencia de las células hijas a permanecer parcialmente adheridas después de la división celular.
Los cocos pueden disponerse: -De a pares y se los llama diplococos. -Si se disponen en cadena se llaman estreptococos. -Cuatro células esféricas conforman una tetrada en forma de racimo o irrregular se llaman estafilococos. -En paquetes cúbicos se denominan sarcinas.
Antes de hablar de lo que es la virologia debemos dar un pequeño concepto de lo que es un virus. La palabra virus viene de veneno. Los agentes que hoy conocemos se designaron hace muchos años con el nombre de “agentes filtrables”. La visualización del virus se realizó en la cristalografía y comprobó la presencia de elementos lineales en series hexagonales que contienen ARN y proteínas. Las enfermedades producidas por virus han afectado a la humanidad desde años remotos. La virología médica está presente en todas las especialidades y áreas médicas. Los virus son partículas formadas por ácido nucleico y proteínas. Considerados como la forma mas pequeña de vida, los virus, son partículas inertes que adquieren vitalidad y se reproducen cuando entran a una célula de un huésped susceptible, y no se multiplican por división, sino que una vez sintetizados sus componentes, estos se ensamblan.
La Virología es la materia que se ocupa del estudio de los virus.Etos son los causantes de numerosas enfermedades.
Los virus son importantes herramientas en investigación. Utilizando virus se ha avanzado en la Biología Molecular, conocimiento de genes, de mecanismos de replicación, trascripción, ác. nucleicos, etc. Los genomas de los virus son muy usados en Ingeniería Genética como vectores para transferir genes de unos individuos a otros.
Tienen importancia biológica. Todos los seres vivos del planeta tienen virus que los parasitan.
MATRICULA:2011-0328 GRUPO:5 CONTINUACION. *ESTRUCTURA GENERAL DE LOS VIRUS.(2)
Como se puede observar en esta imagen todos los virus están constituidos por material genético DNA o RNA (pero no ambos) y una cápside formada por proteínas. Al conjunto del genoma y de la cápside se le denomina núcleo cápside, que puede estar rodeada o no de una envoltura. Podemos diferenciar varios tipos de virus en función de su constitución: desnudos o con envoltura.
*Los virus desnudos, según el tipo de cápside, se diferencian en 3 tipos:
-Icosaédricos. La cápside es un icosaedro.
-Helicoidales. La cápside es un cilindro hueco.
-Complejos. Tienen diferentes estructuras anejas a la cápside (cola,..).
*Los virus con envoltura se diferenciar también en tres tipos:
-Icosaédricos.
-Helicoidales.
-Complejos.
Podemos definir su composición de la siguiente manera:
-Ácidos nucleicos virales:
Los virus se caracterizan, a diferencia de los otros organismos, por presentar una única especie de ácido nucleico constitutiva que puede ser ADN o ARN, monocatenario o bicatenario con estructura de doble hélice.
-Cápsides:
La cápside es una cubierta proteica externa que encierra y protege al genoma viral de la acción de nucleasas y otros factores adversos del medio exterior. Además, en los virus desnudos carentes de envoltura, la cápside es la encargada de establecer a través de alguna de sus proteínas la unión con la célula que será parasitada por el virus. Asimismo, las proteínas de la cápside contienen los determinantes antigénicos contra los que el sistema inmune del huésped elaborará la respuesta de anticuerpos en defensa del organismo.
-Envolturas
La envoltura de un virus es una membrana constituida por una doble capa lipídica asociada a glicoproteínas que pueden proyectarse en forma de espículas desde la superficie de la partícula viral hacia el exterior.
Los virus adquieren su estructura mediante un proceso de brotación a través de alguna membrana celular. El número de glicoproteínas que presentan los virus animales es muy variable.
La simetría es la forma que adopta un virus en el espacio y está dad por la estructura de su nucleocápside. Como se puede observar en la siguiente imagen la simetría puede ser helicoidal, icosaédrica ,binaria o compleja.
-Los virus con simetría icosaédrica son poliedros regulares con 20 caras triangulares, 30 aristas y 12 vértices. Son ejemplos de virus con simetría icosaédrica desnudos el virus de la poliomielitis o los adenovirus. Los herpesvirus y togavirus son virus con simetría icosaédrica y recubiertos por una envoltura.
-Los virus con simetría helicoidal presentan una nucleocápside cilindrica que puede estar extendida como en el virus del mosaico del tabaco. Este es un virus desnudo. En el caso de la imagen que tenemos aquí la nucleocápside está arrollada sobre sí misma y recubierta por una envoltura, como en el virus Influenza.
-los virus con simetría binaria tienen cabeza icosaédrica y cola hélica. El ácido nucleico se encuentra en la cabeza. Bacteriófagos.
-Se denominan virus de simetría compleja a aquellos que presentan una nucleocápside helicoidal o icosaédrica recubierta por una envoltura laxa. Ejemplos de éstos virus son los poxvirus (forma de ladrillo), los rhabdovirus (forma de bala) y algunos bacteriófagos que poseen una cabeza con simetría icosaédrica y una cola con simetría helicoidal.
*TAMAÑO DE LOS VIRUS.(4)
En el siguiente recuadro notamos la variedad de tamaños con respecto a las células que están en comparación.
Se puede decir que los virus son estructuras extraordinariamente pequeñas. La mayoría de los virus poseen un tamaño muy inferior al de las bacterias. Por ello, sólo pueden observarse con el microscopio electrónico.
Dado su pequeño tamaño es necesario utilizar para su medición una medida inferior al micrón, denominada nanómetro (nm) o milimicrón (mu) que equivales a 10-9 metros. Los virus mas pequeños, por ejemplo los que integran la familia Picornaviridae, donde se incluye el virus de la poliomielitis, miden alrededor de 27 nm y los más grandes, como los de la familia Poxviridae, donde se encuentra el virus de la viruela, miden aproximadamente 250 nm.
La mayoría de los virus estudiados tienen un diámetro de entre 10 y 300 nanómetros. Algunos Filovirus tienen un tamaño total de hasta 1.400 nm, sin embargo, sólo miden unos 80 nm de diámetro.La mayoría de virus no pueden ser observados con un microscopio óptico, de manera que se utilizan microscopios electrónicos de barrido y de transmisión para visualizar partículas víricas.
MATRICULA:2011-0328 GRUPO:5 CONTINUACION FAMILIAS DE VIRUS.(5) (patogenos para el hombre)
Aquí se puede notar una gran diversidad de estos entre los cuales podemos mencionar:
-PicoRNAvirus:
Por picornavirus se conoce a todo virus que pertenece a la familia Picornaviridae. Los picornavirus son virus no envueltos, de única cadena ARN con cápside icosaédrica.
Su genoma es inusual ya que tiene una proteína en el extremo 5 ‘que se utiliza como cebador para la transcripción por la ARN polimerasa. El nombre se deriva de “pico”, que significa pequeño, ya que es literalmente un pequeño virus de ARN.
Los picornavirus se separan en una serie de géneros e incluyen muchos importantes patógenos de humanos. Las enfermedades que causan son muy variadas, que van desde aguda “gripe común” a enfermedades como la poliomielitis, y otras infecciones crónicas.
- Los Calicivirus: Son Virus RNA pequeños pertenecientes a la familia Caliciviridae. Dentro de esta familia destacan dos géneros: los Norovirus, también conocidos como "virus tipo Norwalk" o "virus pequeños redondos estructurados" y los Sapporovirus.
- Hepadnavirus: Los hepadnavirus son un subgrupo de los virus que producen hepatitis. Es uno de los virus más importantes desde el punto de vista médico. Infecta a más de un cuarto de billón de personas y es una de las principales causas de cirrosis y cáncer hepático (carcinoma hepatocelular). HBV, también representa el primer ejemplo del éxito de una vacuna recombinante para una enfermedad contagiosa humana, y puede tener importantes implicaciones para otras enfermedades de transmisión sexual, como el HIV.
- Reovirus: Grupo de tres virus de doble cadena de ARN presentes en todas partes, identificados en los aparatos respiratorio y digestivo tanto de personas sanas como de enfermas. Los reovirus han sido implicados en algunos casos de enfermedades de vías respiratorias superiores y de gastroenteritis infantiles.
- Los Rhabdovirus: Son virus con cápside helicoidal. Incluye al virus de la rabia y de la estomatitis vesicular (que es el organismo modelo). El virus de la estomatitis vesicular es un virus de 180x75nm, con cápsida rodeada por una envoltura con proyecciones constituídas por la proteína G.
- Togavirus: Familia de arbovirus a la que pertenecen los organismos causantes de encefalitis, del dengue, de la fiebre amarilla y de la rubéola.
- Coronavirus: Género de virus de la familia de los Coronaviridae El coronavirus humano es el responsable de infecciones respiratorias de las vías superiores. Miembro de la familia de los virus que abarcan distintos tipos capaces de producir una enfermedad respiratoria aguda.
- Bunyavirus: Género de virus ARN de vertebrados de la familia Bunyaviridae.
-Papovavirus: Grupo de pequeños virus ADN, algunos de los cuales pueden ser potencialmente productores de cáncer. La verruga humana está producida por un tipo de papovavirus. Algunos tipos de papovavirus son: papovavirus del papiloma, papovavirus del polioma y papovavirus SV.
- Paramixovirus: Miembro de una familia de virus que abarca a los organismos causantes de la gripe, de las paperas y de algunas infecciones respiratorias.
- Retrovirus: Virus de ácido ribonucleico que posee una enzima que le permite convertir su material genético en ácido desoxirribonucleico cuando ha penetrado en una célula viva. Al grupo de los retrovirus pertenece el virus del sida.
- Filovirus: Género de virus con ácido ribonucleico de cadena simple con sentido negativo, de la familia Filoviridae. El género incluye los virus Ébola y Marburg.
- Poxvirus: Miembro de una familia de virus que incluye a los organismos que producen el molusco contagioso, la viruela y la vacuna.
MATRICULA:2011-0328 GRUPO:5 CONTINUACION *DIAGNOSTICO DE LAS VIROSIS.(6) Antes de hablar de diagnostico es bueno hacernos esta pregunta:
¿Cómo infectan los virus al ser humano? Los virus pueden penetrar en el cuerpo humano por cualquiera de sus “puertas de entrada,” pero por lo general entran por la nariz y la boca. Una vez dentro del cuerpo, el virus se adhiere a la superficie de la célula que piensa atacar, denominada célula huésped. Una vez dentro, comienza a elaborar otros virus idénticos a partir de la proteína de la célula huésped. Los nuevos virus pueden salir a través de la membrana externa de la célula huésped, a la que a veces destruyen antes de abandonarla, y se lanzan al ataque contra otras células huésped. El proceso prosigue hasta que el cuerpo elabora suficiente cantidad de anticuerpos y otras defensas para derrotar a los virus invasores.
-Los síntomas son muy variables y dependen del virus y del órgano afectado. Muchos virus, como muchas bacterias, producen fiebre y síntomas respiratorios (tos, estornudos) o intestinales (náuseas, vómito, diarrea). Las virosis, aun cuando no sean de peligro, a menudo provocan fiebre elevada en los niños de corta edad.
- Algunas infecciones virales, como la gripe, el resfriado comúno, la varicela son fáciles de identificar por sus síntomas, sin que se necesiten análisis de laboratorio. Para otras, como la hepatitis viral, el sida y la mononucleosis infecciosa, se suele analizar una muestra de sangre en busca de anticuerpos específicos contra el virus. La presencia de estos anticuerpos confirma el diagnóstico. - En ciertos casos, los virus pueden cultivarse en tejidos celulares o bien el virus se identifica por su ácido nucleico, valiéndose de una técnica que recibe el nombre de reacción en cadena de la polimerasa (RCP). Las pruebas como el RCP o el cultivo celular se emplean cuando las de anticuerpos no tienen la precisión suficiente o cuando es necesario cuantificar con mayor exactitud el virus invasor.
*ETAPAS DE LA REPLICACION VIRAL.(7)
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
*PENETRACION.(9) Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
-Virus envueltos.( Figura B)
1. Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
2.Entrada vía endosomas en la superficie celular.
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
-Virus no envueltos o desnudos (Figura A)
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
*PENETRACION DE LOS BACTERIOFAGOS.(10)
Los bacteriófagos tienen la modalidad de adsorber y penetrar, mejor conocida por su mecanismo de inyección. Las proteínas que forman las espinas de la placa basal entran en contacto mediante sus receptores con los de la pared bacteriana y se anclan a ella. La accion de algunas enzimas hidroliticas de la capside rompe la pared y esta es perforada por el eje tubular. El genoma se introduce por el eje hasta el citoplasma de la bacteria quedando fuera de la celula la capsida. Los virus vegetales pasan la pared celular a través de pequeñas grietas producidas por picaduras de insectos. Los virus animales tienen acceso mas directo a la membrana de las celulas. Los virus sin envoltura penetran por completo en la célula por endocitosis y una vez dentro de la vesicula,el virus libera enzimas hidroliticas para romper la vesicula, y quedar libre en el citoplasma el virus. Los virus con envoltura penetran al fusionar su cubierta con la membrana celular,y se forma una vesicula endocítica que se une a un endosoma,cuya membrana se fusiona con la envoltura del virus,escapando de ser digerido en su interior.
*REPLICACION Y MADURACION.(11)
La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínas víricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores. Los ribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada. Como podemos observar en la imagen: -Los virus con ADN realizan la replicación del material genético de la misma manera que las células; en el caso de los virus con ADN monocatenario, previamente a la replicación se sintetiza una cadena de ADN complementario para formar la doble hélice. -Los virus con ARN replican el material genético sin necesidad de pasar por ADN, actuando cada cadena de ARN como molde para la síntesis de su complementaria.
MATRICULA:2011-0328 GRUPO:5 CONTINUACION *MADURACION Y SALIDA DE VIRUS DESNUDOS Y ENVUELTOS.(12)
. El desnudamiento ocurre simultáneamente o después de la penetración y comprende la eliminación de todas o parte de las cubiertas proteicas que rodean al ácido nucleico viral, de manera de dejar expuesto dentro de la célula el genoma viral desnudo o asociado a alguna enzima para que pueda expresarse.
En algunos casos sólo se remueve parte de la cápside viral y en esas condiciones el genoma expresa sus funciones. Para otros virus complejos, como por ejemplo los Poxvirus, el desnudamiento parece ocurrir en dos etapas: en la primera hay remoción de ciertas cubiertas proteicas por enzimas celulares, parte del genoma es tanscripto en esas condiciones y luego, en una segunda etapa, y a través de una proteína codificada por el virus, se produce el desnudamiento total del genoma.
En general el desnudamiento es un proceso difícil de estudiar y establecer con precisión sus características por la rapidez con que ocurre y la dificultad en separarlo temporalmente de la penetración. Es por ello que se suele denominar internalización del virión al conjunto de ambos eventos, penetración y desnudamiento.
*SALIDA DE UN VIRUS ENVUELTO.(13)
Vemos aquí como los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes.
No todas las partículas víricas liberadas son infecciosas. La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
Los virus envueltos (como el VIH) son liberados de la célula huésped por gemación. Durante este proceso, el virus adquiere su envoltura, que es una parte modificada de la membrana plasmática del huésped.
MATRICULA:2011-0328 GRUPO:5 CONTINUACION *ANTIVIRALES.(14) Los farmacos que inhiben procesos especificos del virus, como la union a la celula, la decapsidacion del genoma viral o el ensamblaje de los nuevos virus, o, preferentemente, inhiben la sintesis macromolecular dirigida por el virus.
Actualmente se dispone de farmacos antivirales de probada eficacia terapeutica y profilactica para varias infecciones virales comunes y algunas potencialmente mortales. Ademas, continua la busqueda de nuevos farmacos antivirales y nuevas estrategias terapeuticas para el tratamiento de las enfermedades virales.
En el siguiente recuadro se presenta una clasificacion de estos antivirales de la siguiente manera: -Antivirales que impiden la entrada del virus a la celula:
•Amantadina: actua sobre el virus influenza A. Su efecto principal es inhibir la decapsidacion viral o el desensamblaje del virion durante la endocitosis. •Rimantadina: derivado 4-10 veces mas activa que la amantadina. •Oseltamivir: inhibe la actividad de neuraminidasa del virus influenza. •Zanamivir:inhibe la actividad de neuraminidasa del virus influenza. •Docosanol: interfiere en la fusion del VHS-1 y VHS-2 a las membranas de las celulas del huesped. Es de uso topico.
-Antivirales que actuan sobre la fase de replicación del genoma viral.
•Aciclovir: inhibe la sintesis del ADN viral. Tiene accion sobre los herpesvirus. •Valaciclovir: profarmaco del Aciclovir. •Ganciclovir: inhibe la sintesis del ADN viral. Tiene accion sobre los herpesvirus, pero su caracteristica singular es una potente inhibicion de la replicacion del CMV. •Valganciclovir: profarmaco del Ganciclovir. •Adefovir: Actua como inhibidor competitivo de las polimerasas del ADN y transcriptasas inversas virales con respecto al dATP. Se selectividad se relaciona con una mayor afinidad por la ADN polimerasa del VHB que por las polimerasas de las celulas huesped. Esta aprobado para el tratamiento de la hepatitis B cronica. •Cidofovir: inhibe la sintesis de ADN viral de los herpesvirus.
MATRICULA:2011-0328 GRUPO:5 CONTINUACION *MECANISMO DE ACCION DE LOS RETROVIRALES.(15)
Con esta imagen podemos describirlo d ela siguiente manera y tomare como ejemplo el virus del VIH: -Inhibidores de la entrada La infección por el VIH se inicia con la unión de la partícula viral a la membrana externa de las células diana, mediante la interacción entre la gp120 del VIH y dos moléculas celulares: el receptor CD4 y el correceptor de quimocinas (CCR5 o CXCR4). En un paso posterior, la gp41 viral se adhiere a la superficie celular para iniciarse la fusión de las membranas del VIH y de la célula diana. Para que esto ocurra es preciso que la gp41 se pliegue sobre sí misma acortando el espacio que separa ambas membranas. -Inhibidores de transcriptasa inversa Se trata de fármacos que inhiben la síntesis del ADN complementario a partir del ARN viral, función realizada por la transcriptasa inversa (TI) viral en el interior de la célula infectada. Según el mecanismo de bloqueo de la TI se dividen en: -Inhibidores de la integrasa Una vez sintetizado el ADN complementario, éste pasa al interior del núcleo de la célula para allí integrarse en el ADN celular. La integrasa viral es la enzima responsable de este proceso. Actualmente se encuentran en desarrollo clínico una serie de compuestos que bloquean el ciclo viral al nivel de la integración del ADN complementario en el ADN celular. -Inhibidores de la proteasa Como se ha señalado, el ADN complementario viral sintetizado por acción de la TI se integra en el ADN de la célula infectada. El ciclo viral continúa cuando el ADN viral se transcribe a ARN mensajero que en los ribosomas celulares se traduce en una poliproteína viral. Esta cadena proteica ha de escindirse en diversos péptidos virales que dan lugar a las proteínas estructurales y funcionales que formarán parte de las nuevas partículas virales. La proteasa viral es la enzima responsable de la hidrólisis de la poliproteína en los diversos péptidos individuales. Existe un grupo de fármacos, los inhibidores de la proteasa (IP), que basan su mecanismo de acción en el bloqueo de esta enzima. De este modo se impide la maduración y ensamblaje de las partículas virales, y la infección de nuevas células diana.
*RETROVIRALES.(16) Entre las clases de retrovirus que observamos aquí podemos mencionar los siguientes: -Inhibidores nucleosídicos de la transcriptasa inversa. Actúan sobre la enzima transcriptasa inversa, incorporándola a la cadena de ADN que crea el virus. Con ello, esa cadena se vuelve defectuosa e impiden que el virus se reproduzca. Son los siguientes: Zidovudina, abacavir, didanosina, estavudina, lamivudina y tenofovir. -Inhibidores no nucleósidos de la transcriptasa inversa. Bloquean directamente la acción de la enzima y la multiplicación del virus. Son los siguientes: efavirenz, nevirapina y etravirina. -Inhibidores de la proteasa. Actúan sobre la enzima proteasa, bloqueando su acción e impidiendo la producción de nuevas copias de células infectadas por el VIH. Son los siguientes: amprenavir, atazanavir, darunavir, indinavir, lopinavir/r, nelfinavir, ritonavir y saquinavir. -Inhibidores de la fusión. Impiden la entrada de virus a la célula, con lo que no puede reproducirse. La enfuvirtida pertenece a este grupo. -Inhibidores de la integrasa. Bloquean la actividad de la enzima integrasa, responsable de la inserción del ADN del VIH en el ADN humano (código genético de la célula). De esta manera inhibe la replicación del virus y su capacidad de infectar a nuevas células.
MATRICULA:2011-0328 GRUPO:5 CONTINUACION *OTROS BIONES ACELULARES.(20) -Satelites(schneider): Un satélite es un agente subvírico formado por ácido nucleico y que depende de la coinfección de la célula huésped por un virus (helper o máster). Si el satélite codifica una proteína de cubierta en la que se introduce el ácido nucleico, hablamos de virus satélite. Aunque su genoma no guarda relación con el del helper, sí que depende de las enzimas de éste para replicarse. Un ejemplo es el virus satélite del mosaico del tabaco, que utiliza al virus del mosaico del tabaco para llevar a cabo su reproducción - virusoides o sateloides(fransky): aunque son estudiados en virología, no se les considera verdaderos virus sino partículas subvirales, por necesitar de otros virus para propagarse. Por esto, en las clasificaciones de virus aparecen con el término de "ARN satélite circular". De hecho, el término virusoide ha sido también usado genéricamente para referirse a todos los ácidos nucleicos satélites. - Los plásmidos : son moléculas de ADN extracromosómico circular o lineal que se replican y transcriben independientes del ADN cromosómico. Están presentes normalmente en bacterias, y en algunas ocasiones en organismos eucariotas como las levaduras. Su tamaño varía desde 1 a 250 kb. El número de plásmidos puede variar, dependiendo de su tipo, desde una sola copia hasta algunos cientos por célula.
MATRICULA:2011-0328 GRUPO:5 CONTINUACION *PRIONES I .(21) Los priones son los agentes causantes de un grupo de patologías neurodegenerativas letales características de mamíferos, también conocidas como encefalopatías espongiformes transmisibles. Estos agentes son capaces de propagarse dentro de un mismo huesped causando una lesión espongiótica y de transmitirse de huesped a huesped con elevados tiempos de incubación. Los priones se propagan mediante la transmisión de proteínas anómalas con mal plegamiento. Cuando un prion entra en un organismo sano, actúa sobre la forma normal del mismo tipo de proteína existente en el organismo, modificándola y convirtiéndola en prion. Estos priones recién formados pueden pasar a convertir más proteínas, provocando una reacción en cadena que produce grandes cantidades de la proteína prion. Todos los priones conocidos inducen la formación de amiloides plegado, en los que actúan polimerasas formando un agregado que consiste en apretadas hojas ß. El período de incubación de las enfermedades priónicas se determina por la tasa de crecimiento exponencial asociados con la replicación de priones, que es un equilibrio entre el crecimiento lineal y la rotura de los agregados (hay que tener en cuenta que la propagación del prion depende de la presencia de la proteína normalmente plegada en la que los priones pueden inducir plegamiento. Los organismos que no expresan la forma normal de la proteína priónica no pueden desarrollar o transmitir la enfermedad.
*PRIONES II. (22) No se conocen con exactitud los mecanismos a través de los cuales la Prpc pasa a ser PrpSc. Una de las hipótesis es la rotura de los puentes de disulfuro entre aminoácidos de la estructura de hélice alfa (normalmente entre cisteínas que es un tipo de aminoácido que puede formar dichos puentes), hecho que da lugar a radicales –SH, presentes en las láminas beta. Según las hipótesis actuales, los aminoácidos que actúan en el cambio conformacional son la metionina, por su capacidad de cambiar el radical metilo por –H consiguiendo un radical -SH; la cisteína, por su disposición a formar puentes disulfuro; y la valina, por su proximidad atómica a la Met y Cys. En el laboratorio podemos romper los puentes disulfuro mediante dos mecanismos: a partir de la oxidación con ácido perfórmico o mediante la rotura con beta-mercaptoetanol y posterior acetilación con iodoacetato. Otras hipótesis señalan que la sustitución del aminoácido prolina por leucina puede ser el posible responsable de la desestabilización de la estructura terciaria de la proteína y su transformación en agente patógeno, hecho apoyado con la comparación del gen PRNP procendente de un paciente enfermo con el de un paciente sano.
Virología rama de la microbiología y Estudio de los virus y las enfermedades que éstos causan.
Estructura de los virus diap#2
Los virus más pequeños y simples están constituidos únicamente por ácido nucleico y proteínas. El ácido nucleico es el genoma viral, ubicado en el interior de la partícula, y puede ser ADN o ARN.Este conjunto de genoma y proteínas íntimamente asociadas es llamado "core", núcleo, nucleoproteína o nucleoide. Este núcleo central está rodeado por una cubierta proteica, la cápside, que junto con el genoma constituye la nucleocápside. La cápside es una cubierta proteica externa que encierra y protege al genoma viral de la acción de nucleasas y otros factores adversos del medio exterior. La envoltura de un virus es una membrana constituida por una doble capa lipídica asociada a glicoproteínas que pueden proyectarse en forma de espículas desde la superficie de la partícula viral hacia el exterior. Los virus se caracterizan, a diferencia de los otros organismos, por presentar una única especie de ácido nucleico constitutiva que puede ser ADN o ARN, monocatenario o bicatenario con estructura de doble hélice.
La simetría es la forma que adopta un virus en el espacio y está dad por la estructura de su nucleocápside. La simetría puede ser helicoidal, icosaédrica o compleja.
Los virus con simetría helicoidal presentan una nucleocápside cilindrica que puede estar extendida como en el virus del mosaico del tabaco. Este es un virus desnudo. En otros casos la nucleocápside está arrollada sobre sí misma y recubierta por una envoltura, como en el virus Influenza. Los virus con simetría icosaédrica son poliedros regulares con 20 caras triangulares, 30 aristas y 12 vértices. Son ejemplos de virus con simetría icosaédrica desnudos el virus de la poliomielitis o los adenovirus. Los herpesvirus y togavirus son virus con simetría icosaédrica y recubiertos por una envoltura. Se denominan virus de simetría compleja a aquellos que presentan una nucleocápside helicoidal o icosaédrica recubierta por una envoltura laxa. Ejemplos de éstos virus son los poxvirus (forma de ladrillo), los rhabdovirus (forma de bala) y algunos bacteriófagos que poseen una cabeza con simetría icosaédrica y una cola con simetría helicoidal.
Tamaño de los virus diap #4
Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus. pero en realidad los tamaños de los virus se presentan de forma variable.
Familia de virus diap#5
Familia Especies importantes2 Adenoviridae adenovirus Picornaviridae virus de hepatitis A, poliovirus Herpesviridae Virus varicela-zoster Hepadnaviridae virus de hepatitis B Flaviviridae virus de hepatitis C Retroviridae Virus de inmunodeficiencia humana (VIH) Orthomyxoviridae virus de influenza Paramyxoviridae virus virus del sarampión, Papovaviridae Rhabdoviridae virus de la rabia Togaviridae virus de Rubéola
Podemos diagnosticar la virosis mediante metodos. Los métodos utilizados para reconocer las infecciones por virus humanos pueden clasificarse en DIRECTOS e INDIRECTOS
METODOS DIRECTOS
Son aquellos que detectan:
1. El virus como agente infeccioso (aislamiento viral):La base del diagnóstico viral es la detección del virus o de sus componentes. El aislamiento del virus era la técnica standard de oro sobre la cual se medían todas las otras pruebas de diagnóstico viral.
2. La presencia de antígenos virales (técnicas inmunológicas): Inmunofluorescencia (IF), Enzimoinmunoanálisis (EIA), Test de Aglutinación: (!)Esta técnica se puede utilizar para una identificación rápida del virus directamente sobre la muestra. (2) El test de aglutinación ha sido usado para detectar antígeno de Rotavirus en heces
3.La presencia de ácidos nucleicos virales (PCR, etc.).
4.El virus como partícula viral (microscopía electrónica).
Etapas de la replicacion viral diap#7,8,9,10,11,12,13
Dentro de estos podemos mencionar:
1). ADHESION: Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
2).PENETRACIÓN Y PENETRACION DE LOS BACTERIOFAGOS Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. (B) Entada vía endosomas en la superficie celular
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
3). REPLICACION Y MADURACION (MADURACION Y SALIDA DE VIRUS Y ENVUELTOS) El ácido nucleido debe de estar lo suficientemente expuesto como para que la replicación viral comience en esta fase. Cuando el ácido nucleico es expuesto, las partículas víricas infecciosas no pueden removerse de las células – este es el comienz de la fase de ECLIPSE – la cual perdura hasta que viriones infecciosos nuevos sean creados.Nuevas partículas víricas son ensambladas. Puede haber un estadio de maduración posterior al proceso inicial de ensamblaje.
4). SALIDA DE UN VIRUS ENVUELTO Los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes. No todas las partículas víricas liberadas son infecciosas. La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
Antivirales diap#14 Se pueden clasificar en tres grandes grupos a los agentes quimioterapicos activos frente a los virus:
grupo 1: Farmacos Viricidas: farmacos que directamente inactivan los virus intactos: Detergentes, Disolventes organicos: eter, cloroformo, Luz ultravioleta, Podofilinas, las cuales destruyen ademas tejidos del huesped.
Farmacos Antivirales: farmacos que inhiben procesos especificos del virus, como la union a la celula, la decapsidacion del genoma viral o el ensamblaje de los nuevos virus, o, preferentemente, inhiben la sintesis macromolecular dirigida por el virus. Farmacos Inmunomodulares: comprenden los que reemplazan las respuestas inmunitarias deficientes del huesped y los que favorecen a los endogenos.
grupo 2: ANTIVIRALES QUE IMPIDEN LA ENTRADA DEL VIRUS A LA CELULA
Amantadina: actua sobre el virus influenza A. Su efecto principal es inhibir la decapsidacion viral o el desensamblaje del virion durante la endocitosis. Rimantadina: derivado 4-10 veces mas activa que la amantadina. Oseltamivir: inhibe la actividad de neuraminidasa del virus influenza. Zanamivir:inhibe la actividad de neuraminidasa del virus influenza. Docosanol: interfiere en la fusion del VHS-1 y VHS-2 a las membranas de las celulas del huesped.
grupo 3:ANTIVIRALES QUE ACTUAN SOBRE LA FASE DE REPLICACIÓN DEL GENOMA VIRAL
Aciclovir: inhibe la sintesis del ADN viral. Tiene accion sobre los herpesvirus. Valaciclovir: profarmaco del Aciclovir. Ganciclovir: inhibe la sintesis del ADN viral. Tiene accion sobre los herpesvirus, pero su caracteristica singular es una potente inhibicion de la replicacion del CMV. Valganciclovir: profarmaco del Ganciclovir. Adefovir: Actua como inhibidor competitivo de las polimerasas del ADN y transcriptasas inversas virales con respecto al dATP. Se selectividad se relaciona con una mayor afinidad por la ADN polimerasa del VHB que por las polimerasas de las celulas huesped. Esta aprobado para el tratamiento de la hepatitis B cronica. Cidofovir: inhibe la sintesis de ADN viral de los herpesvirus. Penciclovir: inhibe la sintesis de ADN viral de los herpesvirus. Famciclovir. Profarmaco del Penciclovir de administracion oral. Fomivirseno. Inhibe la replicacion del CMV humano. Se administra de forma intravitrea. Foscarnet: actua sobre los herpesvirus y la transcriptasa reversa del HIV. Es inhibidor frente a la mayoria de las cepas de CMV resistentes a ganciclovir y de VHS y VVZ resistentes a aciclovir. Idoxuridina. Inhibe la sintesis de ADN viral. Actua en especial sobre los herpesvirus y poxvirus. Ribavirina: se utiliza en el tratamiento de la infección por HCV y por el VSR. Trifluridina. Vidarabina.
Mecanismo de accion de los retrovirales diap#15 y 16
Los retrovirus son pequeños microorganismos que contienen una única cadena de ARN. Su ciclo vital se basa en la conversión de su ARN geonómico en doble cadena de ADN y posteriormente en la integración de éste último en los cromosomas de la célula huésped en lo que se denomina provirus endógeno. Entre las clases de retrovirus que observamos aquí podemos mencionar los siguientes: -Inhibidores nucleosídicos de la transcriptasa inversa. Actúan sobre la enzima transcriptasa inversa, incorporándola a la cadena de ADN que crea el virus. Con ello, esa cadena se vuelve defectuosa e impiden que el virus se reproduzca. -Inhibidores no nucleósidos de la transcriptasa inversa. Bloquean directamente la acción de la enzima y la multiplicación del virus. -Inhibidores de la proteasa. Actúan sobre la enzima proteasa, bloqueando su acción e impidiendo la producción de nuevas copias de células infectadas por el VIH. -Inhibidores de la fusión. Impiden la entrada de virus a la célula, con lo que no puede reproducirse. -Inhibidores de la integrasa. Bloquean la actividad de la enzima integrasa, responsable de la inserción del ADN del VIH en el ADN humano (código genético de la célula). De esta manera inhibe la replicación del virus y su capacidad de infectar a nuevas células.
MATRICULA: 2011-0546 GRUPO:5
Biones Acelulares diap#17
Los biones celulares son aquellos que se replican. Entre estos encontramos:
Viroides 1 y 2 (#18 y 19): - Pequeñas moléculas de ARN circular de cadena sencilla formadas por unos pocos cientos de nucleótidos. Carecen de cápsida y su tamaño es una milésima parte del de los virus más pequeños. - Aparecen con mayor abundancia en el núcleo de las células infectadas. - Sólo han sido detectados en plantas, en las que producen una gran variedad de enfermedades y pueden ser transmitidos tanto horizontal como verticalmente. - No existen ninguna evidencia de que los viroides son traducidos a proteínas, ni se conoce cómo causan las enfermedades. Lo que sí se sabe es que son replicados por los enzimas del huésped. - Algunas semejanzas entre la secuencia de nucleótidos de los viroides y ciertos secuencias del genoma de las plantas no traducidas (intrones) sugieren que los viroides evolucionaron a partir de los intrones.
Otros biones acelulares (diap#20) Satelites(schneider): Un satélite es un agente subvírico formado por ácido nucleico y que depende de la coinfección de la célula huésped por un virus (helper o máster). Si el satélite codifica una proteína de cubierta en la que se introduce el ácido nucleico, hablamos de virus satélite. Un ejemplo es el virus satélite del mosaico del tabaco, que utiliza al virus del mosaico del tabaco para llevar a cabo su reproducción - virusoides o sateloides(fransky): aunque son estudiados en virología, no se les considera verdaderos virus sino partículas subvirales, por necesitar de otros virus para propagarse. Por esto, en las clasificaciones de virus aparecen con el término de "ARN satélite circular". - Los plásmidos : son moléculas de ADN extracromosómico circular o lineal que se replican y transcriben independientes del ADN cromosómico. Están presentes normalmente en bacterias, y en algunas ocasiones en organismos eucariotas como las levaduras. Su tamaño varía desde 1 a 250 kb. El número de plásmidos puede variar, dependiendo de su tipo, desde una sola copia hasta algunos cientos por célula.
Priones I (diap#21):· Los priones son partículas infecciosas proteicas que provocan las encefalopatías espongiformes transmisibles. · Poseen la misma secuencia aminoacídica de una proteína normal, pero presentan una estructura diferente. · Inducen, por un mecanismo hasta ahora desconocido, la transformación de proteínas normales en anómalas
Priones II (diap#22): Contradicen el dogma central de la biología molecular, según el cual el flujo de información es en todos los seres vivos: ADN ‡ ARNm ‡ Secuencia Aminoácidos ‡ Estructura Tridimensional Proteínas. La teoría de los priones propuesta por Prusiner supone la existencia de dos plegamientos para una única secuencia de amino ácidos. Además, el replegamiento de la PrP normal por acción de la PrP patológica, implica un flujo de información de una proteína a otra a nivel de estructura terciaria
La virología (rama de la microbiologia) es el estudio de los virus: su estructura, clasificación y evolución, su manera de infectar y aprovecharse de las células huésped para la reproducción del virus, su interacción con los organismos huéspedes, su inmunidad, la enfermedad que causan, las técnicas para su aislamiento, cultivo y su uso en investigación y terapia. La virología es considerada un sub-campo de la microbiología y la medicina. La virologia también es una rama taxonómica de la biología para estudiar los virus y todos sus componentes. Su investigación incluye: la replicación viral los patógenos virales. la inmunología viral. las vacunas virales. los métodos de diagnóstico. la quimioterapia antiviral. las medidas de control de una infección. los diferentes signos que manifiestan la presencia de virus.
DIAPOSITIVA # 2: ESTRUCTURA GENERAL DE LOS VIRUS 2011-0226
Como pudimos observar,en esta imagen se presenta la estructura general de los virus,dentro de las cuales encontramos las siguientes:
El ácido nucleico en los virus contiene la información específica y el potencial para modificar operaciones en la célula infectada.68 Los ácidos nucleicos son macromoléculas constituidas por cadenas de nucleótidos, los cuales a su vez están constituidos por una base nitrogenada asociada a un azúcar del grupo de las pentosas y a uno o más grupos de fosfatos. Existen cuatro posibles tipos de ácido nucleico viral:69 ADN de cadena sencilla, ADN de cadena doble, ARN de cadena sencilla y ARN de cadena doble. Virus que contienen cualquiera de estos tipos de ácido nucleico pueden ser encontrados tanto entre los fagos como entre los virus que infectan a plantas o animales. El ADN de algunos bacteriófagos se caracteriza por contener bases raras que substituyen alguna o algunas de las bases normalmente presentes en el ADN.70 El ADN de cadena doble presente en algunos virus, se caracteriza por tener segmentos de cadena sencilla en ambos extremos de la molécula. Debido a que son complementarias las secuencias de nucleótidos presentes en ambos extremos. El ADN naturalmente circular puede ser de cadena sencilla como en el fago ØXI74,70 o de cadena doble, como en el virus SV4O. Existe evidencia de que algunos virus ARN que producen infecciones en vegetales como el limonero y la papa contienen moléculas circulares de ARN.
Los virus presentan una amplia diversidad de formas y tamaños, llamadas «morfologías». Son unas 100 veces más pequeños que las bacterias. La mayoría de los virus estudiados tienen un diámetro de entre 10 y 300 nanómetros. Algunos Filovirus tienen un tamaño total de hasta 1.400 nm, sin embargo, sólo miden unos 80 nm de diámetro. La mayoría de virus no pueden ser observados con un microscopio óptico, de manera que se utilizan microscopios electrónicos de barrido y de transmisión para visualizar partículas víricas.Para aumentar el contraste entre los virus y el trasfondo se utilizan tinciones densas en electrones. Son soluciones de sales de metales pesados como wolframio, que dispersan electrones en las regiones cubiertas por la tinción. Cuando las partículas víricas están cubiertas por la tinción (tinción positiva), oscurecen los detalles finos. La tinción negativa evita este problema, tiñendo únicamente el trasfondo. Una partícula vírica completa, conocida como virión, consiste en un ácido nucleico rodeado por una capa de protección proteica llamada cápside. Las cápsides están compuestas de subunidades proteicas idénticas llamadas capsómeros. Los virus tienen un «envoltorio lipídico» derivado de la membrana celular del huésped. La cápside está formada por proteínas codificadas por el genoma vírico, y su forma es la base de la distinción morfológica.Las subunidades proteicas codificadas por los virus se autoensamblan para formar una cápside, generalmente necesitando la presencia del genoma viral. Sin embargo, los virus complejos codifican proteínas que contribuyen a la construcción de su cápside. Las proteínas asociadas con los ácidos nucleicos son conocidas como nucleoproteínas, y la asociación de proteínas de la cápside vírica con ácidos nucleicos víricos recibe el nombre de nucleocápside.
Envoltura Algunas especies de virus se envuelven en una forma modificada de una de las membranas celulares, o bien es la membrana externa que rodea una célula huésped infectada, o bien membranas internas como la membrana nuclear o el retículo endoplasmático, consiguiendo así una bicapa lipídica exterior conocida como envoltorio vírico. Esta membrana se rellena de proteínas codificadas por el genoma vírico y el del huésped, la membrana lipídica en sí y todos los carbohidratos presentes son codificados completamente por el huésped. El virus de la gripe y el VIH utilizan esta estrategia. La mayoría de virus envueltos dependen de la envoltura para infectar. La cápsida vírica o cápside vírica Es una estructura proteica formada por una serie de monómeros llamados capsómeros. En el interior de esta cápside se encuentra siempre el material genético del virus. Puede estar rodeada por una envoltura. Cada capsómero puede estar constituido por una o varias proteínas distintas. El término nucleocápside se refiere al material genético envuelto en su cápside.
Hay cuatro tipos principales de morfología vírica: Helicoidal las cápsides helicoidales se componen de un único tipo de capsómero apilado alrededor de un eje central para formar una estructura helicoidal que puede tener una cavidad central o un tubo hueco. Esta formación produce viriones en forma de barra o de hilo, pueden ser cortos y muy rígidos, o largos y muy flexibles. El material genético, normalmente ARN monocatenario, pero a veces ADN monocatenario, queda unido a la hélice proteica por interacciones entre el ácido nucleico con carga negativa y la carga positiva de las proteínas. En general, la longitud de una cápside helicoidal está en relación con la longitud del ácido nucleico que contiene, y el diámetro depende del tamaño y la distribución de los capsómeros. El conocido virus del mosaico del tabaco es un ejemplo de virus helicoidal. Icosaédrica la mayoría de virus que infectan los animales son icosaédricos o casi-esféricos con simetría icosaédrica. Un icosaedro regular es la mejor manera de formar una carcasa cerrada a partir de subunidades idénticas. El número mínimo requerido de capsómeros idénticos es doce, cada uno compuesto de cinco subunidades idénticas. Muchos virus, como los rotavirus, tienen más de doce capsómeros y parecen esféricos, manteniendo esta simetría. Los ápices de los capsómeros están rodeados por otros cinco capsómeros y reciben el nombre de pentones. Las caras triangulares de éstos también se componen de otros seis capsómeros y reciben el nombre de hexones. Complejos los virus tienen una cápside que no es ni puramente helicoidal, ni puramente icosaédrica, y que puede poseer estructuras adicionales como colas proteicas o una pared exterior compleja. Algunos bacteriófagos (como el Fago T4) tienen una estructura compleja que consiste en un cuerpo icosaédrico unido a una cola helicoidal (esta cola actúa como una jeringa molecular, atacando e inyectando el genoma del virus a la célula huésped), que puede tener una base hexagonal con fibras caudales proteicas que sobresalgan. Los poxvirus son virus grandes y complejos con una morfología inusual. El genoma vírico está asociado con proteínas dentro de una estructura discal central conocida como nucleoide. El nucleoide está rodeado por una membrana y dos cuerpos laterales de función desconocida. El virus tiene una envoltura exterior con una espesa capa de proteína en la superficie. La partícula en general es ligeramente pleomorfa, con una forma que puede ir de la de un huevo a la de un ladrillo. Mimivirus es uno de los tipos de virus más grandes conocidos, con un diámetro en su cápside de 400 nm. De su superficie se proyectan filamentos proteicos de 100 nm. La cápside, tomada en microscopio electrónico, tiene una forma hexagonal, de manera que probablemente es icosaédrica. Algunos virus que infectan a las Archaeas tienen estructuras inusuales, que no están relacionadas con ningún otro virus conocido. De igual manera, algunos bacteriófagos pueden tener diferentes estructuras en cuanto a su cola, con formas algo raras con respecto a otros virus.
Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus.
Su pequeño tamaño explica lo tardío del descubrimiento de estos agentes. La primera referencia sobre la existencia de los virus se debe al botánico ruso Dimitri Ivanovski en 1892. Este investigador buscaba el agente causante de la enfermedad denominada mosaico del tabaco, y llegó a la conclusión de que debía tratarse de una toxina o de un organismo más pequeño que las bacterias, pues el agente atravesaba los filtros que retenían las bacterias. Denominó a estos agentes patógenos virus filtrables.
En 1897, el microbiólogo holandés Martinus Beijerink realizó experimentos similares a los de Ivanovski, y llegó a desechar la idea de las toxinas, pues se trataba de un agente capaz de reproducirse, ya que mantenía su poder infeccioso de unas plantas a otras, sin diluirse su poder patógeno. Poco después, los microbiólogos alemanes Frederick Loeffler y Paul Frosch descubrieron que la fiebre aftosa del ganado era producida por un virus filtrable que actuaba como un agente infeccioso.
En la década de los 30, con el uso de filtros de tamaño de poro inferior, con las técnicas de cultivo celular in vitro que permitían la obtención de gran cantidad de virus, con la ultracentrifugación y finalmente con el microscopio electrónico y la difracción de rayos X, se logró visualizar a estos agentes.
En esta imagen podemos observar las familias de virus patogenos para el hombre,dentro de los cuales podemos desctacar:
- Bunyavirus: Género de virus ARN de vertebrados de la familia Bunyaviridae.
-Papovavirus: Grupo de pequeños virus ADN, algunos de los cuales pueden ser potencialmente productores de cáncer. La verruga humana está producida por un tipo de papovavirus. Algunos tipos de papovavirus son: papovavirus del papiloma, papovavirus del polioma y papovavirus SV.
- Paramixovirus: Miembro de una familia de virus que abarca a los organismos causantes de la gripe, de las paperas y de algunas infecciones respiratorias.
- Retrovirus: Virus de ácido ribonucleico que posee una enzima que le permite convertir su material genético en ácido desoxirribonucleico cuando ha penetrado en una célula viva. Al grupo de los retrovirus pertenece el virus del sida.
- Filovirus: Género de virus con ácido ribonucleico de cadena simple con sentido negativo, de la familia Filoviridae. El género incluye los virus Ébola y Marburg.
- Poxvirus: Miembro de una familia de virus que incluye a los organismos que producen el molusco contagioso, la viruela y la vacuna.
- Reovirus: Grupo de tres virus de doble cadena de ARN presentes en todas partes, identificados en los aparatos respiratorio y digestivo tanto de personas sanas como de enfermas. Los reovirus han sido implicados en algunos casos de enfermedades de vías respiratorias superiores y de gastroenteritis infantiles.
- Los Rhabdovirus: Son virus con cápside helicoidal. Incluye al virus de la rabia y de la estomatitis vesicular (que es el organismo modelo). El virus de la estomatitis vesicular es un virus de 180x75nm, con cápsida rodeada por una envoltura con proyecciones constituídas por la proteína G.
- Togavirus: Familia de arbovirus a la que pertenecen los organismos causantes de encefalitis, del dengue, de la fiebre amarilla y de la rubéola.
- Coronavirus: Género de virus de la familia de los Coronaviridae El coronavirus humano es el responsable de infecciones respiratorias de las vías superiores. Miembro de la familia de los virus que abarcan distintos tipos capaces de producir una enfermedad respiratoria aguda.
- Los Calicivirus: Son Virus RNA pequeños pertenecientes a la familia Caliciviridae. Dentro de esta familia destacan dos géneros: los Norovirus, también conocidos como "virus tipo Norwalk" o "virus pequeños redondos estructurados" y los Sapporovirus.
- Hepadnavirus: Los hepadnavirus son un subgrupo de los virus que producen hepatitis. Es uno de los virus más importantes desde el punto de vista médico. Infecta a más de un cuarto de billón de personas y es una de las principales causas de cirrosis y cáncer hepático (carcinoma hepatocelular). HBV, también representa el primer ejemplo del éxito de una vacuna recombinante para una enfermedad contagiosa humana, y puede tener importantes implicaciones para otras enfermedades de transmisión sexual, como el HIV.
-PicoRNAvirus:
Por picornavirus se conoce a todo virus que pertenece a la familia Picornaviridae. Los picornavirus son virus no envueltos, de única cadena ARN con cápside icosaédrica.
Su genoma es inusual ya que tiene una proteína en el extremo 5 ‘que se utiliza como cebador para la transcripción por la ARN polimerasa. El nombre se deriva de “pico”, que significa pequeño, ya que es literalmente un pequeño virus de ARN.
Los picornavirus se separan en una serie de géneros e incluyen muchos importantes patógenos de humanos. Las enfermedades que causan son muy variadas, que van desde aguda “gripe común” a enfermedades como la poliomielitis, y otras infecciones crónicas.
DIAPOSITIVA #6: DIAGNOSTICO DE LA VIROSIS 2011-0226
Los métodos utilizados para reconocer las infecciones por virus humanos pueden clasificarse en DIRECTOS e INDIRECTOS, según persigan demostrar la presencia del virus o de alguno de sus constituyentes (antígeno o genoma viral) o bien la respuesta de anticuerpos específicos por parte del huésped en el curso de la infección.
Gran parte de las técnicas utilizadas en el diagnóstico clínico se basan en pruebas serológicas que identifican anticuerpos específicos frente a diversas proteínas antigénicas. Sin embargo, existen circunstancias en las cuales son necesarias pruebas que detecten precozmente la infección viral (tratamientos específicos, medidas profilácticas, etc.).
En algunas infecciones virales es posible detectar la presencia de antígenos virales previamente al desarrollo de la seroconversión, siendo esta prueba la única evidencia de la exposición al virus cuando no existe aumento de los niveles de anticuerpos circulantes (pacientes inmunodeprimidos).
Igualmente la detección del genoma viral puede favorecer la precocidad del diagnóstico viral y su confirmación. En la última década se han desarrollado una serie de técnicas para el diagnóstico viral basadas en la detección de ácidos nucleicos. De ellas la Reacción en cadena de la polimerasa (PCR) es la más utilizada.
En el momento actual, la tendencia en el diagnóstico virológico consiste en emplear nuevas y más sensibles tecnologías de detección de antígenos y de investigación de ácidos nucleicos con el propósito de lograr un diagnóstico viral más rápido. El desarrollo de quimioterapia antiviral efectiva es responsable de esta tendencia que hace de la identificación rápida, sensible y específica de los virus, una necesidad.
PRINCIPALES TÉCNICAS METODOS DIRECTOS Son aquellos que detectan: 1. El virus como agente infeccioso (aislamiento viral). 2. La presencia de antígenos virales (técnicas inmunológicas): Inmunofluorescencia (IF), Enzimoinmunoanálisis (EIA), Test de Aglutinación. 3.La presencia de ácidos nucleicos virales (PCR, etc.). 4.El virus como partícula viral (microscopía electrónica).
DIAPOSITIVA# 7 : ETAPAS DE LA REPLICACION VIRAL 2011-0226
En lo que respecta a este tema podemos ver que los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
DIAPOSITIVA#8 : ADHESION 2011-0226
La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado.
Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre las proteínas de la cápsida. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica. Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras: Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma. Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma. Los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos: Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma. Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
DIAPOSITIVA #10: PENETRANCIA DE LOS BACTERIOGRAFOS 2011-0226
El bacteriófago está formado por un corpúsculo que contiene ácido nucleico y una cola con la extremidad en placa mediante la que se fija sobre la bacteria y la infecta. Los bacteriófagos tienen la propiedad de lisar ciertos microbios (colibacilo, bacilos tífico y paratífico, bacilo disentérico, estafilococo, etc) en los que penetra y se multiplica: éstos son los bacteriófagos virulentos que desencadenan cada vez la lisis bacteriana. Los bacteriófagos atenuados pueden, bajo forma de profagos, permanecer inactivos dentro de las bacterias que infectan durante largo tiempo (bacterias lisógenas). Los bacteriófagos tienen la modalidad de adsorber y penetrar, mejor conocida por su mecanismo de inyección. Las proteínas que forman las espinas de la placa basal entran en contacto mediante sus receptores con los de la pared bacteriana y se anclan a ella. La accion de algunas enzimas hidroliticas de la capside rompe la pared y esta es perforada por el eje tubular. El genoma se introduce por el eje hasta el citoplasma de la bacteria quedando fuera de la celula la capsida. Los virus vegetales pasan la pared celular a través de pequeñas grietas producidas por picaduras de insectos. Los virus animales tienen acceso mas directo a la membrana de las celulas. Los virus sin envoltura penetran por completo en la célula por endocitosis y una vez dentro de la vesicula,el virus libera enzimas hidroliticas para romper la vesicula, y quedar libre en el citoplasma el virus. Los virus con envoltura penetran al fusionar su cubierta con la membrana celular,y se forma una vesicula endocítica que se une a un endosoma,cuya membrana se fusiona con la envoltura del virus,escapando de ser digerido en su interior
DIAPOSITIVA#11: REPLICACION Y MADURACION 2011-0226
La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínas víricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores. Los ribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada. Los virus con ADN realizan la replicación del material genético de la misma manera que las células; en el caso de los virus con ADN monocatenario, previamente a la replicación se sintetiza una cadena de ADN complementario para formar la doble hélice. Los virus con ARN replican el material genético sin necesidad de pasar por ADN, actuando cada cadena de ARN como molde para la síntesis de su complementaria. Los retrovirus constituyen una excepción a lo dicho anteriormente, ya que su ARN sintetiza un ADN bicatenario, que será el que posteriormente realice la síntesis de nuevos ejemplares de RNA vírico. Los virus con ADN y los retrovirus sintetizan el ARN a partir de la cadena molde de ADN de forma similar a como lo hacen las células. Los virus con ARN, excepto los retrovirus, sintetiza en el ARN copiando la cadena molde de ARN, sin necesidad de pasar por ADN. Posteriormente a estos procesos, tiene lugar el ensamblaje de las piezas para construir nuevos viriones. En muchos virus, como el del mosaico del tabaco, el ensamblaje es automático y depende de la concentración salina del medio. En otros virus, en el ensamblaje intervienen enzimas codificadas en el ácido nucleico del virus.
DIAPOSITIVA #12: MADURACION Y SALIDA DE VIRUS DESNUDOS Y ENVUELTOS 2011-0226
En esta diapositiva podemos ver un esquema muy detallado de la maduracion y salida de virus desnudos y envueltos. El desnudamiento ocurre simultáneamente o después de la penetración y comprende la eliminación de todas o parte de las cubiertas proteicas que rodean al ácido nucleico viral, de manera de dejar expuesto dentro de la célula el genoma viral desnudo o asociado a alguna enzima para que pueda expresarse.
En algunos casos sólo se remueve parte de la cápside viral y en esas condiciones el genoma expresa sus funciones. Para otros virus complejos, como por ejemplo los Poxvirus, el desnudamiento parece ocurrir en dos etapas: en la primera hay remoción de ciertas cubiertas proteicas por enzimas celulares, parte del genoma es tanscripto en esas condiciones y luego, en una segunda etapa, y a través de una proteína codificada por el virus, se produce el desnudamiento total del genoma.
En general el desnudamiento es un proceso difícil de estudiar y establecer con precisión sus características por la rapidez con que ocurre y la dificultad en separarlo temporalmente de la penetración. Es por ello que se suele denominar internalización del virión al conjunto de ambos eventos, penetración y desnudamiento.
DIAPOSITIVA#13: SALIDA DE UN VIRUS ENVUELTO 2011-0226
Aqui podemos ver claramente la forma en que sale un virus envuelto;los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes.
No todas las partículas víricas liberadas son infecciosas. La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
Los virus envueltos (como el VIH) son liberados de la célula huésped por gemación. Durante este proceso, el virus adquiere su envoltura, que es una parte modificada de la membrana plasmática del huésped.
Los antivirales son un tipo de medicamento usado para el tratamiento de infecciones producidas por virus. Tal como los antibióticos (específicos para bacteria), existen antivirales específicos para distintos tipos de virus. No sin excepciones, son relativamente inocuos para el huésped, por lo que su aplicación es relativamente segura. Deben distinguirse de los viricidas, que son compuestos químicos que destruyen las partículas virales presentes en el medio ambiente. Muchos de los antivirales disponibles actualmente están diseñados para ayudar el tratamiento del VIH (virus del sida), herpesvirus, productores de la varicela, el herpes labial, el herpes genital, etc. y los virus de la hepatitis B y C, que pueden causar cáncer de hígado. Los investigadores están trabajando actualmente para extender el rango de antivirales a otras familias de patógenos. Para que un fármaco antiviral sea considerado para el tratamiento de una infección viral debe cumplir ciertos requisitos indispensables. El fármaco debe ser específicamente activo contra el virus “target” (blanco), inhibiendo algún paso esencial de su metabolismo y debe poder debilitar a las cepas resistentes que puedan surgir. Además, debe tener ciertos parámetros que permitan al fármaco ser de alta biodisponibilidad oral, rápida penetración en los tejidos infectados y atóxicos para las células normales y el organismo en general a corto y largo plazo. El fármaco debe ser transformado a su forma activa solamente en células infectadas, debe ser excretado sin ser metabolizado o sin generar metabolitos tóxicos, y debe tener baja frecuencia de administración. Finalmente, el fármaco debe tener bajo costo para que pueda estar disponible para toda la población. Todas estas características, entre otras, dificultan el descubrimiento de nuevos fármacos efectivos para el tratamiento de las enfermedades virales.
DIAPOSITIVA #15: MECANISMOS DE ACCION DE LOS ANTIVIRALES 2011-0226
Los fármacos antirretrovirales no matan al virus de la inmunodeficiencia humana (VIH). Ni siquiera logran dormirlo. Lo 'atontan' un poco y lo mantienen en un estado de reposo, pero que le permite, de vez en cuando, salir a 'pasear' por las células del organismo humano. Lo ha descubierto un equipo de investigadores españoles, que pone así fin a una de las controversias médicas más extendidas en el campo del sida.
Durante años, la teoría más apoyada ha sido la que consideraba que el cóctel de fármacos impedía la replicación del virus. No era capaz de eliminarlo, pero sí podía evitar que siguiera infectando. Otra hipótesis, más minoritaria, indicaba que a pesar de la terapia, el virus podía mantener algo de actividad. Y es esta última la que finalmente se ha confirmado, según la investigación que publica 'Nature Medicine' y que ha sido realizada por expertos del Instituto de Investigación del Sida IrsiCaixa, ubicado en el Hospital German Trias i Pujol de Badalona.
Lo que han visto los investigadores, dirigidos por Javier Martínez Picado, es que algunos pacientes infectados por el VIH, a pesar de que presentan una carga viral indetectable gracias al tratamiento y que parecen tener la infección controlada, cuentan con una pequeña proporción del virus que sigue siendo infeccioso. Un descubrimiento que, según sus autores, "tiene importantes implicaciones clínicas, ya que podría explicar por qué, a pesar de que los pacientes permanecen en terapia durante muchos años, los fármacos no permiten curar la infección de manera definitiva".
Martínez Picado lo aclara de forma sencilla a ELMUNDO.es: "Un paciente infectado sin tratamiento es como un bosque en llamas. Con los fármacos logramos apagar el fuego, pero quedan rescoldos y basta con un pequeño golpe de aire para que vuelva a incendiar. Esto es lo que hace el virus. Está apagado, pero no muerto".
Dentro de los retrovirales podemos encontrar los siguientes grupos: -Inhibidores nucleosídicos de la transcriptasa inversa. Actúan sobre la enzima transcriptasa inversa, incorporándola a la cadena de ADN que crea el virus. Con ello, esa cadena se vuelve defectuosa e impiden que el virus se reproduzca. Son los siguientes: Zidovudina, abacavir, didanosina, estavudina, lamivudina y tenofovir. -Inhibidores no nucleósidos de la transcriptasa inversa. Bloquean directamente la acción de la enzima y la multiplicación del virus. Son los siguientes: efavirenz, nevirapina y etravirina. -Inhibidores de la proteasa. Actúan sobre la enzima proteasa, bloqueando su acción e impidiendo la producción de nuevas copias de células infectadas por el VIH. Son los siguientes: amprenavir, atazanavir, darunavir, indinavir, lopinavir/r, nelfinavir, ritonavir y saquinavir. -Inhibidores de la fusión. Impiden la entrada de virus a la célula, con lo que no puede reproducirse. La enfuvirtida pertenece a este grupo. -Inhibidores de la integrasa. Bloquean la actividad de la enzima integrasa, responsable de la inserción del ADN del VIH en el ADN humano (código genético de la célula). De esta manera inhibe la replicación del virus y su capacidad de infectar a nuevas células.
DIAPOSITIVA#17,18 Y 19: BIONES ACELULARES 2011-0226
Aqui podemos encontrar a los viroides 1 y 2. - Pequeñas moléculas de ARN circular de cadena sencilla formadas por unos pocos cientos de nucleótidos. Carecen de cápsida y su tamaño es una milésima parte del de los virus más pequeños. - Aparecen con mayor abundancia en el núcleo de las células infectadas. - Sólo han sido detectados en plantas, en las que producen una gran variedad de enfermedades y pueden ser transmitidos tanto horizontal como verticalmente. - No existen ninguna evidencia de que los viroides son traducidos a proteínas, ni se conoce cómo causan las enfermedades. Lo que sí se sabe es que son replicados por los enzimas del huésped. - Algunas semejanzas entre la secuencia de nucleótidos de los viroides y ciertos secuencias del genoma de las plantas no traducidas (intrones) sugieren que los viroides evolucionaron a partir de los intrones.
DIAPOSITIVA #20: OTROS BIONES ACELULARES 2011-0226
AQUI PODEMOS ENCONTRAR LO SIGUIENTE: -Satelites(schneider): Un satélite es un agente subvírico formado por ácido nucleico y que depende de la coinfección de la célula huésped por un virus (helper o máster). Si el satélite codifica una proteína de cubierta en la que se introduce el ácido nucleico, hablamos de virus satélite. Aunque su genoma no guarda relación con el del helper, sí que depende de las enzimas de éste para replicarse. Un ejemplo es el virus satélite del mosaico del tabaco, que utiliza al virus del mosaico del tabaco para llevar a cabo su reproducción - virusoides o sateloides(fransky): aunque son estudiados en virología, no se les considera verdaderos virus sino partículas subvirales, por necesitar de otros virus para propagarse. Por esto, en las clasificaciones de virus aparecen con el término de "ARN satélite circular". De hecho, el término virusoide ha sido también usado genéricamente para referirse a todos los ácidos nucleicos satélites. - Los plásmidos : son moléculas de ADN extracromosómico circular o lineal que se replican y transcriben independientes del ADN cromosómico. Están presentes normalmente en bacterias, y en algunas ocasiones en organismos eucariotas como las levaduras. Su tamaño varía desde 1 a 250 kb. El número de plásmidos puede variar, dependiendo de su tipo, desde una sola copia hasta algunos cientos por célula.
DIAPOSITIVA #21: PRIONES 2011-0226
Un prion1 es un tipo de proteína patógena (es decir, una molécula compleja que induce enfermedades), más específicamente una psialoproteína patógena, la cual tiene alterada su estructura secundaria, teniendo un incorrecto plegamiento de su estructura terciaria.2 A diferencia del resto de los agentes infecciosos (virus, bacterias, hongos etc...), que contienen ácidos nucleicos (ya sea ADN, el ARN, o ambos), un prion sólo esta compuesto por aminoácidos y no presenta material genético. El prion, palabra acuñada en 1982 por Stanley B. Prusiner al investigar una serie de enfermedades de carácter crónico e irreversibles que afectaban al sistema nervioso central,3 es un acrónimo inglés derivado de las palabras proteína e infección. Los priones son los responsables de las encefalopatías espongiformes transmisibles en una variedad de mamíferos, incluida la encefalopatía espongiforme bovina (EEB, también conocida como "enfermedad de las vacas locas") en el ganado y la enfermedad de Creutzfeldt-Jakob (ECJ) en humanos.4 Dichas proteínas mutadas forman agregados supramoleculares y son patógenas con plegamientos anómalos ricos en láminas beta, y autoreproducibles. Los priones se propagan mediante la transmisión de proteínas anómalas con mal plegamiento. Cuando un prion entra en un organismo sano, actúa sobre la forma normal del mismo tipo de proteína existente en el organismo, modificándola y convirtiéndola en prion. Estos priones recién formados pueden pasar a convertir más proteínas, provocando una reacción en cadena que produce grandes cantidades de la proteína prion. Todos los priones conocidos inducen la formación de amiloides plegado, en los que actúan polimerasas formando un agregado que consiste en apretadas hojas ß. El período de incubación de las enfermedades priónicas se determina por la tasa de crecimiento exponencial asociados con la replicación de priones, que es un equilibrio entre el crecimiento lineal y la rotura de los agregados (hay que tener en cuenta que la propagación del prion depende de la presencia de la proteína normalmente plegada en la que los priones pueden inducir plegamiento. Los organismos que no expresan la forma normal de la proteína priónica no pueden desarrollar o transmitir la enfermedad5 .
No se conocen con exactitud los mecanismos a través de los cuales la Prpc pasa a ser PrpSc. Una de las hipótesis es la rotura de los puentes de disulfuro entre aminoácidos de la estructura de hélice alfa (normalmente entre cisteínas que es un tipo de aminoácido que puede formar dichos puentes), hecho que da lugar a radicales –SH, presentes en las láminas beta. Según las hipótesis actuales, los aminoácidos que actúan en el cambio conformacional son la metionina, por su capacidad de cambiar el radical metilo por –H consiguiendo un radical -SH; la cisteína, por su disposición a formar puentes disulfuro; y la valina, por su proximidad atómica a la Met y Cys. En el laboratorio podemos romper los puentes disulfuro mediante dos mecanismos: a partir de la oxidación con ácido perfórmico o mediante la rotura con beta-mercaptoetanol y posterior acetilación con iodoacetato. Otras hipótesis señalan que la sustitución del aminoácido prolina por leucina puede ser el posible responsable de la desestabilización de la estructura terciaria de la proteína y su transformación en agente patógeno, hecho apoyado con la comparación del gen PRNP procendente de un paciente enfermo con el de un paciente sano.
#1 Virología Virología es el estudio de virus y virus como agentes: clasificación y evolución, su estructura, sus formas de infectar y explotar las células para la reproducción del virus, las enfermedades que causan, las técnicas para aislar y la cultura de ellos y su uso en la investigación y terapia.Virología es considerada una parte de la microbiología o de patología. Una importante rama de la Virología es la clasificación de virus. Virus pueden clasificarse según la célula huésped que infectan: virus animales, virus de plantas, hongos virus y bacteriófagos (virus que infectan bacterias, que son los virus más complejos). Otra clasificación utiliza la forma geométrica de su cápside (a menudo una hélice o un icosaedro) o la estructura del virus (por ejemplo, la presencia o la ausencia de un envolvente de lípidos). Gama de virus en tamaño de unos 30 nm a aproximadamente 450 nm, lo que significa que la mayoría de ellos no puede verse con microscopios de luces.
mat:2011-0054 #2 Estructura general de los virus Los virus más pequeños y simples están constituidos únicamente por ácido nucleico y proteínas. El ácido nucleico es el genoma viral, ubicado en el interior de la partícula, y puede ser ADN o ARN. Generalmente está asociado con un número pequeño de moléculas proteicas que pueden tener actividad enzimática o cumplir alguna función estabilizadora para el plegamiento del ácido nucleico y armado de la partícula viral. Este conjunto de genoma y proteínas íntimamente asociadas es llamado "core", núcleo, nucleoproteína, onucleoide. Este núcleo central está rodeado por una cubierta proteica, la cápside, que junto con el genoma constituye la nucleocápside. Las cápsides virales están formadas por un gran número de subunidades polipeptídicas que se ensamblan adoptando una simetría de tipo helicoidal (nucleocápside en forma de bastón) o icosaédrica (partículas casi esféricas). En algunos virus más complejos, por fuera de la cápside se encuentra otra cubierta, la envoltura, que es una estructura membranosa constituida por lípidos y glicoproteínas. Dicha cubierta viral puede ser considerada una cubierta protectora adicional. Los virus se caracterizan, a diferencia de los otros organismos, por presentar una única especie de ácido nucleico constitutiva que puede ser ADN o ARN, monocatenario o bicatenario con estructura de doble hélice. La cápside es una cubierta proteica externa que encierra y protege al genoma viral de la acción de nucleasas y otros factores adversos del medio exterior. Además, en los virus desnudos carentes de envoltura, la cápside es la encargada de establecer a través de alguna de sus proteínas la unión con la célula que será parasitada por el virus. Asimismo, las proteínas de la cápside contienen los determinantes antigénicos contra los que el sistema inmune del huésped elaborará la respuesta de anticuerpos en defensa del organismo. La envoltura de un virus es una membrana constituida por una doble capa lipídica asociada a glicoproteínas que pueden proyectarse en forma de espículas desde la superficie de la partícula viral hacia el exterior. Los virus adquieren su estructura mediante un proceso de brotación a través de alguna membrana celular. El número de glicoproteínas que presentan los virus animales es muy variable.
mat:2011-0054 #3 Simetría virales El conocimiento de la estructura viral es necesario para comprender la interacción de las partículas virales con los receptores de la superficie celular y con los anticuerpos neutralizantes. Será la base para idear medicamentos específicos capaces de bloquear la adherencia del virus, el desprendimiento de su envoltura o las dos cosas en las células susceptibles.
La simetría es la forma que adopta un virus en el espacio y está dad por la estructura de su nucleocápside. La simetría puede ser helicoidal, icosaédrica o compleja. Los virus con simetría helicoidal presentan una nucleocápside cilindrica que puede estar extendida como en el virus del mosaico del tabaco. Este es un virus desnudo. En otros casos la nucleocápside está arrollada sobre sí misma y recubierta por una envoltura, como en el virusInfluenza. Toda la simetría cúbica observada en los virus de los animales, hasta la fecha, es del patrón icosaédrico, la disposición más eficiente de subunidades en una cubierta cerrada. Conocer las reglas que orientan la simetría icosaédrica hace posible determinar el número de capsómeros en una partícula, característica importante de la clasificación de los virus. El ácido nucleico viral se condensa dentro de las partículas isoméricas; en la condensación del ácido nucleico en una forma adecuada para su acumulación participan proteínas "nucleares" codificadas por el virus o, en el caso de los papovirus, histonas celulares. No han podido definirse aún las reglas para la incorporación del ácido nucleico en las partículas isométricas; probablemente una "secuencia de empaque" participa en el ensamble, aunque en general no son cruciales las estructuras primarias, secundarias y terciarias del ácido nucleico. Existen limitaciones de tamaño en las moléculas de ácido nucleico que se pueden empacar o acumular en una cápside icosaédrica determinada. Las cápsides icosaédricas se forman de manera independiente del ácido nucleico. La mayor parte de los preparados de virus isométricos contendrán algunas partículas "vacías", desprovistas de ácido nucleico viral. Ejemplos de simetría cúbica son los grupos de virus tanto de ADN como de ARN. Los virus con simetría icosaédrica son poliedros regulares con 20 caras triangulares, 30 aristas y 12 vértices. Son ejemplos de virus con simetría icosaédrica desnudos el virus de la poliomielitis o losadenovirus. Los herpesvirus y togavirus son virus con simetría icosaédrica y recubiertos por una envoltura. En los casos de simetría helicoidal, las subunidades proteínicas están fijas de manera periódica al ácido nucleico viral, con lo que lo amplían en una espiral. El complejo filamentoso de proteína y ácido nucleico viral (nucleocápside) se enrolla a continuación dentro de una cubierta que contiene lípidos. Por lo tanto, a diferencia del caso de las estructuras icosaédricas, existe una interacción periódica regular entre la proteína de la cápside y el ácido nucleico en el virus que tiene simetría helicoidal. No es posible que se formen partículas helicoidales "vacías". Todos los ejemplos conocidos de los virus de animales con simetría helicoidal contienen genomas de ARN y, con excepción de los rabdovirus, cuentan con nucleocápsides flexibles que están enrolladas en una esfera dentro de las cubiertas. Se denominan virus de simetría compleja a aquellos que presentan una nucleocápside helicoidal o icosaédrica recubierta por una envoltura laxa. Ejemplos de éstos virus son los poxvirus (forma de ladrillo), los rhabdovirus (forma de bala) y algunos bacteriófagos que poseen una cabeza con simetría icosaédrica y una cola con simetría helicoidal. Algunas partículas virales no presentan simetría cúbica o helicoidal simple, sino que tienen una estructura más complicada. Por ejemplo, los poxvirus tienen forma de ladrillos con rebordes sobre su superficie exteARN, y un centro o núcleo y cuerpos laterales en su interior
mat:2011-0054 #4 Tamaño de los virus Aquí se explica el tamaño que poseen los diferentes virus, La mayoría de los virus poseen un tamaño muy inferior al de las bacterias. Por ello, sólo pueden observarse con el microscopio electrónico. Dado su pequeño tamaño es necesario utilizar para su medición una medida inferior al micrón, denominada nanómetro (nm) o milimicrón (mu) que equivales a 10-9 metros. Los virus mas pequeños, por ejemplo los que integran la familia Picornaviridae, donde se incluye el virus de la poliomielitis, miden alrededor de 27 nm y los más grandes, como los de la familiaPoxviridae, donde se encuentra el virus de la viruela, miden aproximadamente 250 nm. Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus.
mat:2011-0054 #5 Familias de virus Inicialmente después de que los virus fueran descubiertos no había sistema para clasificar virus. Por Lo Tanto los virus fueron nombrados casual. La mayor parte de los virus vertebrados se han nombrado según: las enfermedades asociadas (virus de la polio, rabia) el tipo de enfermedad causado (virus murine de la leucemia), los sitios en el cuerpo afectado o de cuál fue aislado el virus primero (rinovirus, adenovirus) los lugares de donde primero fueron aislados (virus de Sendai, Coxsackievirus) los científicos que los descubrieron (virus de Epstein-Barr), o debido a la influencia cultural común del ` de la gripe de las opiniones e.g' del espíritu maligno malo del ` del aire o de la dengue' Según la clasificación, los virus se agrupan según sus propiedades, no las células que infectan. Las consideraciones principales eran el tipo de DNA nucléica del ácido o ARN. Cuatro características debían ser utilizadas para la clasificación de todos los virus: Tipo del ácido nucléico incluyendo la talla del genoma, el strandedness (simple o doble), el lineal o circular, el positivo o el negativo (sentido), segmentos (número y talla), serie y G+C Etc. contento. Simetría del shell de la proteína Presencia o ausencia de una membrana del lípido Dimensiones o la talla del virion y del capsid Otras propiedades incluyen las propiedades fisicoquímicas incluyendo la masa molecular, pH, estabilidad térmica, susceptibilidad a las substancias químicas y a los extremos físicos y al éter y a los detersorios. Según la clasificación, los virus se agrupan según sus propiedades, no las células que infectan. Las consideraciones principales eran el tipo de DNA nucléica del ácido o ARN. Cuatro características debían ser utilizadas para la clasificación de todos los virus: Tipo del ácido nucléico incluyendo la talla del genoma, el strandedness (simple o doble), el lineal o circular, el positivo o el negativo (sentido), segmentos (número y talla), serie y G+C Etc. contento. Simetría del shell de la proteína Presencia o ausencia de una membrana del lípido Dimensiones o la talla del virion y del capsid Otras propiedades incluyen las propiedades fisicoquímicas incluyendo la masa molecular, pH, estabilidad térmica, susceptibilidad a las substancias químicas y a los extremos físicos y al éter y a los detersorios
mat:2011-0054 #6 Diagnostico de los viroides Actualmente se han caracterizado 200 especies de viroides que infectan solamente plantas superiores. Los viroides pueden infectar monocotiledóneas y dicotiledóneas, y tanto especies leñosas como herbáceas. La gama de huéspedes de los viroides es muy variable. Las enfermedades más conocidas causadas por viroides en plantas (y de mayor transcendencia económica) son:Cadang-Cadang del cocotero; Exocortis de los cítricos; Piel de manzana marcada por cicatrices; enfermedad del aguacate tostado/quemado por el sol, el atrofiamiento del tomate, la enfermedad del tubérculo filiforme de la patata y La enfermedad de las patatas ahusadas. A diferencia de los virus vegetales, los viroides se replican, acumulan y expresan sus síntomas mucho más eficientemente a temperaturas elevadas (30-33 °C) (Sänger y Ramon, 1975) y a intensidades de luz asimismo altas. Los viroides -al igual que los priones- son lo máximo en simplicidad y han sido descubiertos hace relativamente poco tiempo. Constituidos por moléculas de ARN desnudo se encuentran en las plantas y se replican en células susceptibles. La molécula puede ser líneal (dando lugar a una estructura en horquilla) o circular (de forma aplanada); en todo caso, los pares de bases complementarios se unen dando como resultado la estructura de doble cadena, semejante a la del ADN. El primer viroide caracterizado, conocido como PSTVd, es una molécula de ARN de cadena simple que contiene 359 nucleótidos. Los viroides no causan enfermedades en los humanos ni en los animales; sus efectos se limitan a las plantas. Sin embargo, un agente infeccioso similar, un viruosoide, causa la hepatitis D, una variedad excepcionalmente virulenta de esa enfermedad. Los virusoides son moléculas de ARN de una sola hebra, como los viroides, pero requieren un virus asistente para infectar. En el caso de la hepatitis D, el viruosoide necesita de la cubierta proteica del virus de la hepatitis B para transmitirse.
mat:2011-0054 #7 Etapas de la replicación viral El ciclo de replicación de los virus consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden sobrevivir por si mismos en el exterior. Para reproducirse se introducirán en células donde controlarán sus mecanismos reproductivos. Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada. La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
mat:2011-0054 #8 Adhesión El primer paso en la infección viral es la absorción a la membrana de la célula susceptible por medio de la adhesión de ligandos virales (proteínas de la capside o glucoproteínas de las espículas, por ejemplo a receptores superficiales de la célula). La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus. Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor. La mayoría de los fagos se adsorben (pegan) a la pared celular de las bacterias susceptibles, pero algunos fagos también son capaces de adsorberse a los flagelos, vellosidades (pili) o cápsulas presentes en la superficie de la bacteria hospedera. El caso mejor documentado de adsorción viral está representado por los fagos T2 y T4, los cuales son virus con morfología compleja que incluye una cabeza, cola, placa basal, clavijas y fibras de la cola. La pegada inicial de estos fagos a los receptores presentes en la superficie de la bacteria ocurre por medio de los extremos distales de las fibras de la cola. Estas fibras largas que hacen la primera unión se doblan por en medio, de manera que sus extremos distales hacen contacto con la pared celular a muy corta distancia del punto medio de la partícula viral. Después de ocurrida la adhesión, la partícula viral se aproxima a la superficie de la célula. Cuando la placa basal del fago se encuentra a unos 100Å (angstrom= 10-10m) de la pared celular, se establece contacto entre las pequeñas clavijas de la placa basal y la pared celular, pero no existe evidencia de que la propia placa basal se adhiera a la pared celular
mat:2011-0054 #9 Penetracion Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre lasproteínas de la cápsida. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica. El caso mejor estudiado de la penetración de un virus en la célula hospedera está representado por el caso del fago T2. La cola de este fago es contráctil y en su forma extendida consiste de 24 anillos de subunidades que forman una funda que rodea a un elemento central. Cada anillo consta de 6 subunidades pequeñas y 6 subunidades mayores. Después de la adsorción del fago a la pared celular, ocurre una contracción de la cola que resulta en una fusión de las subunidades pequeñas y grandes para dar 12 anillos de 12 subunidades. El núcleo de la cola no es contráctil y por lo tanto es expulsado e impulsado a través de las capas externas de la bacteria; a continuación, la cabeza del fago se contrae y esto resulta en la inyección del ADN viral en la célula bacteriana. Este proceso posiblemente es facilitado por la presencia de la enzima lisozima en la cola del fago; esta enzima es capaz de digerir las proteínas de las cubiertas bacterianas; además, hay 144 moléculas de adenosina trifosfato (ATP) en la funda de la cola del fago; la energía para la contracción de esta funda proviene de la conversión de la ATP en adenosina difosfato (ADP) por medio de una reacción hidrolítica que libera un grupo fosfato de la ATP Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras: Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma. Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma. Los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos: Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma. Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
mat:2011-0054 #10 Penetración de los bacteriófagos Muchos bacteriófagos carecen de fundas contráctiles y todavía se desconoce el mecanismo detallado por medio del cual penetran en las bacterias. Sin embargo, existe evidencia de que algunos de estos fagos introducen en la célula material proteico además del ácido nucleico. Las células animales carecen de pared celular y sólo están rodeadas por la membrana plasmática que es muy flexible y cambiante en sus componentes estructurales. Las células animales continuamente introducen elementos del medio externo por medio del proceso de pinocitosis y a través de un procedimiento similar, pero inverso, exportan al medio diversas substancias como enzimas, hormonas y neurotransmisores. Los virus animales solamente pueden infectar células que poseen receptores específicos para el virus en particular. Por lo tanto, se requieren diferentes receptores para diferentes tipos de virus. Se conoce con poco detalle el mecanismo preciso por medio del cual los virus penetran en las células animales. Buena parte del problema radica en que la mayoría de las partículas virales que infectan a una célula son incapaces de iniciar con éxito la multiplicación del virus. Los virus con envoltura pueden penetrar a la célula hospedera por medio de la fusión entre las envolturas virales y la membrana de la célula. Tanto los virus con envoltura como los virus que carecen de ésta pueden ser introducidos a la célula por medio del proceso de pinocitosis (figura III.3.). La fusión de membranas ocasiona la liberación del genoma viral en el citoplasma de la célula, pero en el caso de la pinocitosis el virus entero es contenido en una vesícula formada a partir de la membrana plasmática. Es probable que esta vesícula se fusione a su vez con alguna de las membranas internas de la célula y la cubierta viral es modificada a consecuencia de esta fusión, lo que da lugar a la liberación del ácido nucleico viral. Es importante hacer notar que la penetración del virus y la subsecuente eliminación de las envolturas virales no implican forzosamente la presencia intracelular de ácido nucleico viral desnudo. El ácido nucleico viral puede permanecer unido a las proteínas internas del virus, las cuales pueden incluir enzimas necesarias para la replicación del virus. En el caso de cierto tipo de virus ARN en los cuales el genoma viral sirve directamente como ARN mensajero (el ácido nucleico a partir del cual se sintetizan las proteínas), el ARN viral se asocia con los ribosomas de la célula. Esto ejemplifica el hecho de que el ácido nucleico viral nunca permanece extendido como un verdadero filamento dentro de la célúla, sino que se enrolla y asocia con proteínas de manera que alcanza un estado favorable de mínima energía libre.
mat:2011-0054 #11 Replicación y Maduración Cuando se han sintetizado un número crítico de varios componentes virales, éstos se ensamblan en partículas víricas maduras en el núcleo y/o citoplasma de la célula infectada. En este ensamblaje y al contrario de lo que ocurre en los bacteriófagos, no están implicados enzimas biosintéticos especiales, sino que ocurre de forma espontánea debido a la gran afinidad de la cápsida con el ácido nucleico viral. La maduración se puede definir como la fase de la infección viral durante la cual los componentes estructurales del virión se producen y ensamblan con el nuevo ácido nucleico viral para formar la nucleocápsida. La maduración viral es a veces un proceso ineficaz. Pueden acumularse cantidades excesivas de componentes virales y participar en la formación de cuerpos de inclusión en la célula (ver antes). Como resultado de los profundos efectos dañinos de la replicación viral, se desarrollan por último efectos citopáticos celulares y la célula muere. Sin embargo, hay casos en que el virus no lesiona a la célula y se producen infecciones persistentes y prolongadas REPLICACION DE VIRUS DNA: Después de la adsorción, penetración y eliminación de la cubierta, el ácido nucleico viral es liberado y migra al núcleo de la célula hospedadora. Posteriormente empieza a transcribirse la porción de DNA vírico correspondiente a los genes tempranos. Tiene lugar la traducción, siendo los productos de estos genes enzimas requeridos para la multiplicación del DNA vírico. En la mayoría de los virus DNA, la transcripción temprana se lleva a cabo con la transcriptasa (RNA polimerasa) del hospedador, salvo en el caso de los poxvirus. Algún tiempo después de la iniciación de la multiplicación del DNA, comienza la transcripción y traducción de los genes tardíos que todavía no habían empezado a expresarse. La síntesis de las proteínas de la cápsida ocurre en el citoplasma de la célula hospedadora. Posteriormente las proteínas de la cápsida migran al núcleo donde tiene lugar la maduración: el DNA vírico y las proteínas de la cápsida se ensamblan para formar las partículas víricas completas.
REPLICACION DE VIRUS RNA: La multiplicación de los virus RNA es esencialmente la misma que la de los virus DNA excepto que los mecanismos para generar el mRNA son distintos en los diferentes grupos de virus RNA. Una vez que el RNA vírico y las proteínas son sintetizados, la maduración tiene lugar de una manera similar en el citoplasma. A diferencia de lo que sucede con el ADN, la replicación del ARN es un fenómeno único de los virus ya que la maquinaria genética de la célula se basa exclusivamente en el uso del ADN como templado (ADN-ADN o ADN-ARN). Por lo tanto, para la duplicación de su genoma, así como acontecía para la síntesis de sus ARNm, los virus ARN deben necesariamente aportar la enzima ARN polimerasa ARN dependiente.
mat:2011-0054 #12 Maduración y salida de virus desnudos y envueltos La liberación de los viriones maduros es el paso final en la multiplicación viral. Los mecanismos de liberación varían según el tipo de virus. En algunos virus la célula hospedadora se desintegra liberando los viriones. También pueden ser liberados a ciertos períodos de tiempo mediante exocitosis o a través de canales especiales (túbulos). El número de partículas víricas que se obtienen varía según el tipo de virus, tipo de célula y las condiciones de crecimiento. El rendimiento medio en plantas y animales va desde varios miles a cerca de un millón de viriones por célula, comparado con los varios cientos de fagos que se obtienen en una célula bacteriana. En las etapas tardías del ciclo de multiplicación viral las moléculas de ácido nucleico y proteínas virales sintetizadas en la célula infectada deben ensamblarse para formar los nuevos viriones y salir al espacio extracelular para estar en condiciones de infectar nuevas células. Este proceso de maduración y ensamble, también denominado morfogénesis, comprende el armado de la cápside a partir de las unidades estructurales y luego la formación del virión completo. El mecanismo va a diferir en base a dos propiedades del virión: la estructura de la cápside y la presencia o no de envoltura. El desnudamiento ocurre simultáneamente o después de la penetración y comprende la eliminación de todas o parte de las cubiertas proteicas que rodean al ácido nucleico viral, de manera de dejar expuesto dentro de la célula el genoma viral desnudo o asociado a alguna enzima para que pueda expresarse. En algunos casos sólo se remueve parte de la cápside viral y en esas condiciones el genoma expresa sus funciones. Para otros virus complejos, como por ejemplo los Poxvirus, el desnudamiento parece ocurrir en dos etapas: en la primera hay remoción de ciertas cubiertas proteicas por enzimas celulares, parte del genoma es tanscripto en esas condiciones y luego, en una segunda etapa, y a través de una proteína codificada por el virus, se produce el desnudamiento total del genoma. En general el desnudamiento es un proceso difícil de estudiar y establecer con precisión sus características por la rapidez con que ocurre y la dificultad en separarlo temporalmente de la penetración. Es por ello que se suele denominar internalización del virión al conjunto de ambos eventos, penetración y desnudamiento.
mat:2011-0054 #13 Salida de un virus envuelto Virus que tiene una envoltura o capa exterior. Esta envoltura proviene de la célula infectada o del huésped, en un proceso llamado “gemación”. Durante el proceso de gemación, las partículas víricas recién formadas se “envuelven” o cubren con una capa externa formada por una pieza pequeña de la membrana plasmática de la célula. La envoltura puede cumplir la función de ayudar a un virus a sobrevivir e infectar a otras células.
Los genomas virales recién sintetizados y los polipéptidos de la cápside se ensamblan para formar los virus descendientes. Las cápsides icosaédricas pueden condensarse en ausencia de ácido nucleico, en tanto que las nucleocápsides de los virus de simetría helicoidal no pueden formarse sin ARN viral. No existen mecanismos especiales para la liberación de virus carentes de cubierta; las células infectadas acaban por experimentar lisis y liberar las partículas virales. Los virus cubiertos maduran por un proceso de gemación. Se insertan glucoproteínas de cubierta específica del virus en las membranas celulares y, a continuación, las nucleocápsides hacen gemación a través de la membrana a nivel de estos sitios modificados y, al hacerlo así, adquieren su cubierta. La gemación ocurre a menudo a nivel de la membrana plasmática y puede abarcar a otras membranas de la célula. Los virus cubiertos no son infecciosos hasta que han adquirido sus cubiertas. Por lo tanto, no se acumulan de manera típica viriones descendientes infecciosos en el interior de la célula infectada. Dado que la envoltura está constituida por lipoproteínas, todos los virus con envoltura son muy sensibles a los solventes de lípidos, tales como éter y sales biliares. En la envoltura se encuentran glicoproteínas que constituyen importantes antígenos del virus. Las funciones de la envoltura son similares a las de la cápside. Así, en los virus envueltos, es la envoltura la que presenta las glicoproteínas que permitirán la unión a receptores de la célula huésped. Todos los virus con envoltura son muy sensibles a la desecación, al pH ácido, a los solventes de lípidos y a las sales biliares. Esto hace que su transmisión fecal-oral no sea posible.
En cambio, la mayoría de los virus desnudos son resistentes al medio externo, a la desecación y a los solventes de lípidos. Por esta razón, los virus desnudos pueden penetrar por vía digestiva ya que resisten el pH ácido del estómago. Pueden ser transmitidos por vía fecal.oral, a través de agua o alimentoscontaminados o por manos u objetos. Son ejemplos de virus desnudos el Poliovirus, y losAdenovirus.
mat:2011-0054 #14 Antivirales Los antivirales son un tipo de medicamento usado para el tratamiento de infecciones producidas por virus. Tal como los antibióticos (específicos para bacteria), existen antivirales específicos para distintos tipos de virus. No sin excepciones, son relativamente inocuos para el huésped, por lo que su aplicación es relativamente segura. Deben distinguirse de los viricidas, que son compuestos químicos que destruyen las partículas virales presentes en el medio ambiente.Actualmente se dispone de farmacos antivirales de probada eficacia terapeutica y profilactica para varias infecciones virales comunes y algunas potencialmente mortales. Ademas, continua la busqueda de nuevos farmacos antivirales y nuevas estrategias terapeuticas para el tratamiento de las enfermedades virales. Farmacos que inhiben procesos especificos del virus, como la union a la celula, la decapsidacion del genoma viral o el ensamblaje de los nuevos virus, o, preferentemente, inhiben la sintesis macromolecular dirigida por el virus. Para que un fármaco antiviral sea considerado para el tratamiento de una infección viral debe cumplir ciertos requisitos indispensables. El fármaco debe ser específicamente activo contra el virus“target” (blanco), inhibiendo algún paso esencial de su metabolismo y debe poder debilitar a las cepas resistentes que puedan surgir. Además, debe tener ciertos parámetros que permitan al fármaco ser de alta biodisponibilidad oral, rápida penetración en los tejidos infectados y atóxicos para las células normales y el organismo en general a corto y largo plazo. El fármaco debe ser transformado a su forma activa solamente en células infectadas, debe ser excretado sin ser metabolizado o sin generar metabolitos tóxicos, y debe tener baja frecuencia de administración. Finalmente, el fármaco debe tener bajo costo para que pueda estar disponible para toda la población. Todas estas características, entre otras, dificultan el descubrimiento de nuevos fármacos efectivos para el tratamiento de las enfermedades virales.
ANTIVIRALES QUE IMPIDEN LA ENTRADA DEL VIRUS A LA CELULA
Amantadina: actua sobre el virus influenza A. Su efecto principal es inhibir la decapsidacion viral o el desensamblaje del virion durante la endocitosis. Rimantadina: derivado 4-10 veces mas activa que la amantadina. Oseltamivir: inhibe la actividad de neuraminidasa del virus influenza. Zanamivir:inhibe la actividad de neuraminidasa del virus influenza. Docosanol: interfiere en la fusion del VHS-1 y VHS-2 a las membranas de las celulas del huesped. Es de uso topico.
ANTIVIRALES QUE ACTUAN SOBRE LA FASE DE REPLICACIÓN DEL GENOMA VIRAL
Aciclovir: inhibe la sintesis del ADN viral. Tiene accion sobre los herpesvirus. Valaciclovir: profarmaco del Aciclovir. Ganciclovir: inhibe la sintesis del ADN viral. Tiene accion sobre los herpesvirus, pero su caracteristica singular es una potente inhibicion de la replicacion del CMV. Valganciclovir: profarmaco del Ganciclovir. Adefovir: Actua como inhibidor competitivo de las polimerasas del ADN y transcriptasas inversas virales con respecto al dATP. Se selectividad se relaciona con una mayor afinidad por la ADN polimerasa del VHB que por las polimerasas de las celulas huesped. Esta aprobado para el tratamiento de la hepatitis B cronica. Cidofovir: inhibe la sintesis de ADN viral de los herpesvirus. Penciclovir: inhibe la sintesis de ADN viral de los herpesvirus. Famciclovir. Profarmaco del Penciclovir de administracion oral. Fomivirseno. Inhibe la replicacion del CMV humano. Se administra de forma intravitrea. Foscarnet: actua sobre los herpesvirus y la transcriptasa reversa del HIV. Es inhibidor frente a la mayoria de las cepas de CMV resistentes a ganciclovir y de VHS y VVZ resistentes a aciclovir. Idoxuridina. Inhibe la sintesis de ADN viral. Actua en especial sobre los herpesvirus y poxvirus. Ribavirina: se utiliza en el tratamiento de la infeccion por HCV y por el VSR. Trifluridina. Vidarabina.
mat:2011-0054 #15 Mecanismos de acción de los retrovirales
Los retrovirus son pequeños microorganismos que contienen una única cadena de ARN . Su ciclo vital se basa en la conversión de su ARN geonómico en doble cadena de ADN y posteriormente en la integración de éste último en los cromosomas de la célula huésped en lo que se denomina provirus endógeno. De forma ocasional algunas secuencias celulares son recombinadas con el provirus y a su vez estas recombinaciones pueden transferirse a otra población celular infectada, lo que puede tener importantes consecuencias en las funciones celulares. Inhibidores de transcriptas inversa: Se trata de fármacos que inhiben la síntesis del ADN complementario a partir del ARN viral, función realizada por la transcriptasa inversa (TI) viral en el interior de la célula infectada. Según el mecanismo de bloqueo de la TI se dividen en: Inhibidores competitivos (ICTI) análogos de nucleósidos/nucleótidos:Se trata de moléculas de estructura similar a los nucleótidos naturales que, al incorporarse como nucleótido anómalo al centro catalítico de la TI, bloquean y finalizan la síntesis del ADN complementario viral. Para que estos fármacos tengan actividad antiviral es preciso que se fosforilen en el interior de las células humanas, mediante la acción de kinasas celulares. Inhibidores no competitivos (INCTI) no-análogos de nucleósidos: Estos fármacos se unen a la TI en un lugar alejado del centro catalítico, donde se incorporan los nucleótidos naturales. El mecanismo de inhibición enzimática de estos compuestos se basa en la modificación de la estructura terciaria de la TI, lo que finalmente se traduce en una menor actividad procesiva3. Se han comercializado dos fármacos de esta familia (nevirapina [NVP] y efavirenz [EFV]). Inhibidores de la integrasa: Una vez sintetizado el ADN complementario, éste pasa al interior del núcleo de la célula para allí integrarse en el ADN celular. La integrasa viral es la enzima responsable de este proceso. Actualmente se encuentran en desarrollo clínico una serie de compuestos que bloquean el ciclo viral al nivel de la integración del ADN complementario en el ADN celular. Inhibidores de la proteasa: Como se ha señalado, el ADN complementario viral sintetizado por acción de la TI se integra en el ADN de la célula infectada. El ciclo viral continúa cuando el ADN viral setranscribe a ARN mensajero que en los ribosomas celulares se traduce en una poliproteína viral. Esta cadena proteica ha de escindirse en diversos péptidos virales que dan lugar a las proteínas estructurales y funcionales que formarán parte de las nuevas partículas virales. La proteasa viral es la enzima responsable de la hidrólisis de la poliproteína en los diversos péptidos individuales. Existe un grupo de fármacos, los inhibidores de la proteasa (IP), que basan su mecanismo de acción en el bloqueo de esta enzima. De este modo se impide la maduración y ensamblaje de las partículas virales, y la infección de nuevas células diana.
mat:2011-0054 #16 Retrovirales Inhibidores de transcriptasa inversa: La transcriptasa inversa es una enzima viral que en el caso del VIH, este requiere para su reproducción. La inhibición de esta enzima impide el desarrollo de ADN viral basado en su ARN. Existen tres clases:
Inhibidores análogos nucleósidos Fueron la primera clase de antirretrovirales desarrollados. Zidovudina: Llamada anteriormente azidotimidina, sus siglas son AZT o ZDV y está comercilaizado como Retrovir® por GlaxoSmithKline (GSK); es un análogo de la timidina
Didanosina: También conocida por su sigla ddI, nombre comercial: Videx®, Videx EC®, análogo de la adenosina.
Zalcitabina: También conocida por su sigla ddC, dideoxicitidina, nombre comercial: Hivid®, análogo de pirimidinaTercer antirretroviral aprobado por la FDA.
Estavudina: También llamada d4T, Nombre comercial: Zerit®, Zerit XR®, análogo de timidina Cuarto antirretroviral aprobado por la FDA el 24 de junio de 1994 y en 1996 para uso pediátrico..
Lamivudina: también llamada 3TC, nombre comercial : Epivir®, análogo de citidina..
Abacavir: también llamado ABC, nombre comercial : Ziagen®, análogo de guanosina.
Emtricitabina: también llamada FTC, nombre comercial : Emtriva®, (anteriormente Coviracil), análogo de citidina,aprobado por la FDA el 2 de julio de 2003.
Festinavir: también llamado BMS-986001. Se trata de un análogo de timidina "de segunda generación" similar a d4T pero sin sus problemas de toxicidad.
EFdA: (4'-etinil-2-fluoro-2'-deoxiadenosina) es un compuesto en desarrollo propiedad de Merck inicialmente desarrollado por la biotecnológica japonesa Yasama Corporation y que ha sido estudiado con el apoyo de amFAR y el NIH de EEUU.
Inhibidores de transcriptasa no nucleósidos Nevirapina: nombre comercial : Viramune®primero de esta clase aprobado por la FDA el 21 de junio de 1996 en adultos y en 1998 para uso pediátrico. Aprobado en España en 1999. Comercializado por Boehringer-Ingelheim.
Delavirdina: nombre comercial : Rescriptor®aprobado por la FDA el 4 de abril de 1997. Actualmente no es de uso coriente debido a sus efectos secundarios adversos. No aprobado para su uso en Europa.
Efavirenz: nombres comerciales: Sustiva®, y Stocrin®; aprobado por la FDA el 21 de septiembre de 1998.
Etravirina: nombre comercial: Intelence®comercializado en España por Janssen-Cilag. Aprobado en Europa para su uso en pacientes previamente tratados.
Rilpivirina: Nombre comercial Edurant®. Rilpivirina se ha comparado con Efavirenz y he demostrado tener menos efectos secundarios a nivel del sistema nervioso central, pero por otro lado, hubo más fracasos virológicos al tratamiento entre los pacientes que tomaron Rilpivirina frente a los que tomaron Efavirenz, especialmente en los pacientes con cargas virales iniciales de más de 100.000 copias/mL.
MK-1439: Es un nuevo inhibidor de la transcriptasa no nucleósido de Merck, que actualmente está en la fase II/III de desarrollo clinico
Inhibidores de proteasa La proteasa es una enzima requerida por el VIH para el ensamblaje final de los viriones, por lo cual su inhibición resulta en inhibición de la replicación viral. Saquinavir, comercializado como Invirase® por Roche. Ritonavir, comercializado como Norvir®, por Abbott. Actualmente se utiliza combinado con todos los inhibidores de proteasa a dosis pequeñas (100 o 200 mg) como potenciador (no como antiviral), ya que es un potente inhibidor del citocromo P450, lo que hace que los niveles en sangre de los inhibidores de proteasa se mantenga más tiempo en cifras óptimas. Es un inhibidor de la proteasa con muchos efectos adversos, pero ha de darse a dosis baja con todos los inhibidores de proteasa para que éstos sean efectivos, la única excpeción es el atazanavir. Indinavir, comercializado como Crixivan® por Merck. Nelfinavir, comercializado como Viracept® por Roche. Amprenavir, nombre comercial: Agenerase® aprobado el 15 de abril de 1999. Fue comercializado por GlaxoSmithKline.Requería de la toma de ocho cápsulas de tamaño muy grande dos veces al día, por lo cual la producción fue finalizada el 31 de diciembre de 2004.
Lopinavir, que éstá comercializado como Kaletra®, si bien incluye también bajas dosis de ritonavir (Kaletra es una combinación a dosis fija de lopinavir y ritonavir). Un inconveniente del Kaletra es que debía conservarse a temperaturas no superiores a 25 ºC, lo que es una dificultad en muchos países en vías de desarrollo. Atazanavir: nombre comercial : Reyataz® aprobado el 20 de junio de 2003. Comercializado por Bristol-Myers Squibb. Primer inhibidor de la proteasa aprobado para 1 toma al día. Parece estar menos asociado con la lipodistrofia y la dislipidemia. Parece no tener resistencia cruzada con otros inhibidores de proteasa. Fosamprenavir, nombre comercial: Lexiva® (EEUU) o Telzir® (Europa) pro-fármaco de amprenavir, aprobado el 20 de octubre de 2003. En España está disponible desde 2005. Comercializado por GlaxoSmithKline. En el organismo es metabolizado para formar amprenavir, el cual es el ingrediente activo. Es en realidad una versión de amprenavir de liberación lenta, lo cual reduce la cantidad de comprimidos necesarios. Debe administrarse siempre con ritonavir como potenciador. Tipranavir o Aptivus® su nombre comercial. Comercializado por Boehringer-Ingelheim. Nuevo inhibidor de la proteasa, que es efectivo contra virus que son resistentes a otros ::inhibidores de la proteasa. Actualmente se utiliza poco. Debe administrarse siempre con ritonavir. Darunavir o Prezista® su nombre comercial. Efectivo contra virus resistentes a otros inhibidores de proteasa. Se utiliza principalmente como componente de tratamiento de rescate, ya que su elevado precio hace que no sea coste-efectivo utiizarlo para iniciar tratamiento. Debe tomarse siempre potenciado con ritonavir
Inhibidores de transcriptasa análogos de nucleótido Normalmente, los análogos nucleósidos se convierten en el cuerpo a nucleótidos, por lo cual el saltarse este paso deriva en menor toxicidad y disminuye el tiempo de latencia del fármaco. Tenofovir disoproxil fumarato: o tenofovir o TDF, nombre comercial: Viread® Aprobado el 26 de octubre de 2001 en EEUU. tenofovir alafenamida: o TAF. Es un nuevo profármaco de tenofovir, en desarrollo y propiedad de Gilead Sciences. De administración una vez al día. Esta nueva formulcación hace que haya menos tenofovir libre, por lo que se espera que disminuyan los efectos secundarios asociados a TDF, especialmente la toxicidad renal. CMX 157: es un profármaco de tenofovir en desarrollo y propiedad de Merck. Además de las ventajas de mejor distribución del fármaco activo aportadas por TAF, CMX 157 tiene una farmacocinética que permitiría la administración 1 vez a la semana. Inhibidores de la integrasa Medicamentos que evitan la acción de la integrasa, selectivos únicamente al VIH-1. Son potentes antirretrovirales y se ha demostrado cierta eficacia en regímenes sin análogos de nucleósidos, pero siempre en combinación (pej Inhibidor de Proteasa + Inhibidor de integrasa). Los regímenes de tratamiento con inhibidores de integrasa producen una recuperación de CD4 más rápida que otros regímenes. Este hecho, si bien es positivo, no está claro si tiene algún beneficio clínico claro. Raltegravir o Isentress® su nombre comercial. Se administra vía oral dos pastillas al día. Disponible en España desde 2008. Está en desarrollo una nueva formulación de administración una vez al día. Elvitegravir , de Gilead Sciences. No está disponible como fármaco independiente. Se administra vía oral una sola pastilla al día. Requiere ser administrado con ritonavir como potenciador, por lo que no estaría libre de los efectos adversos de éste. Se metaboliza por el citocromo P450, por lo que puede tener interferencias medicamentosas con fármacos que se metabolizan por esta vía. Dolutegravir , de ViiV Healthcare. Actualmente en vías de desarrollo (actualmente en fase II-III). Se administra una vez al día y no requiere ser coadministrado con ritonavir ni con ningún otro potenciador. Se metaboliza por la vía del citorcromo P450, con el consiguiente riesgo de interacciones medicamentosas
mat:2011-0054 #17 Biones acelulares Estos microorganismos se clasifican en biones celulares y estos se reproducen y se divide en procariotas y eucariotas y biones acelulares que se replican, por ejemplo los virus, que tienen ácidos nucleicos y los sin ácidos nucleicos que estan los priones. El término acelular significa literalmente "sin células". Se aplica principalmente a los virus, pero también a provirus y viroides, que son entidades biológicas no formadas por células, a diferencia de bacterias, compuestas por células procariotas, y de hongos, vegetales y animales, formados por células eucariotas.
mat:2011-0054 #18 Viroides 1 Los viroides son agentes infecciosos que, al igual que los virus, tienen un ciclo extracelular que se caracteriza por la inactividad metabólica y unciclo intracelular en el que causan infección al huésped susceptible, pero que a diferencia de los virus, los viroides no poseen proteínas ni lípidos y están constituidos por una cadena cíclica corta de ARN, circular o con forma de varilla, (que no codifica proteínas).1 Es importante decir que tanto su forma intracelular como extracelular son las mismas (ARN desnudo), los mecanismos por los cuales éstos logran causar infección están relacionados con la autocatálisis de su material genético. En sí constituyen una etapa primitiva de los virus. El primer viroide fue descubierto por T. O. Diener en 1978, al intentar identificar el agente causal de una enfermedad que inicialmente supuso era inducida por un virus llamada "la enfermedad del tubérculo fusiforme de la patata" (potato spindle tuber o PSTVd) La nomenclatura de los viroides incluye el sufijo “Vd” para distinguirlos de los virus. El segundo viroide en ser descubierto fue el agente causal de la exocortis de los cítricos (citrus exocortis viroid, CEVd). Actualmente se conocen más de 200 viroides.
mat:2011-0054 #19 Viroides 2 Los viroides son los agentes infecciosos de menor complejidad genética y estructural conocidos y representan una forma extrema de parasitismo. Están constituidos exclusivamente por moléculas de ARN de cadena simple, cerradas covalentemente o con forma de bastón de bajo peso molecular (246 a 1994 nucleótidos). Carecen de actividad de ARN mensajero y se replican de forma autónoma, utilizando el sistema de transcripción de la célula suceptible.2 Los viroides se encuentran, casi exclusivamente, en el núcleo de las células infectadas; se desconoce el modo en que se replican pero se sabe que el ARN que los constituyen no funciona como ARN mensajero y tampoco se traduce a enzimas que participen en su propia replicación. Los viroides no codifican para ninguna proteína y llevan a cabo su replicación en la célula hospedadora a través de la RNA polimerasa II codificada por la célula hospedadora aunque también se ha propuesto la posibilidad de que lo haga gracias a una RNA polimerasa dependiente de RNA presente en algunas células eucariotas (Cann, 2001). Dada su localización, se presume que causan la enfermedad interfiriendo con la regulación génica de la célula hospedadora en la etapa de corrección del ARN mensajero celular: particularmente en la eliminación de intrones y de empalmes de exones, presentando secuencias muy similares a los intrones que normalmente son cortados. Sus ARN se replican varias veces. Actualmente se han caracterizado 200 especies de viroides que infectan solamente plantas superiores. Los viroides pueden infectar monocotiledóneas y dicotiledóneas, y tanto especies leñosas como herbáceas. La gama de huéspedes de los viroides es muy variable. Las enfermedades más conocidas causadas por viroides en plantas (y de mayor transcendencia económica) son:Cadang-Cadang del cocotero; Exocortis de los cítricos; Piel de manzana marcada por cicatrices; enfermedad del aguacate tostado/quemado por el sol, el atrofiamiento del tomate, la enfermedad del tubérculo filiforme de la patata y La enfermedad de las patatas ahusadas. A diferencia de los virus vegetales, los viroides se replican, acumulan y expresan sus síntomas mucho más eficientemente a temperaturas elevadas (30-33 °C) (Sänger y Ramon, 1975) y a intensidades de luz asimismo altas. Los viroides -al igual que los priones- son lo máximo en simplicidad y han sido descubiertos hace relativamente poco tiempo. Constituidos por moléculas de ARN desnudo se encuentran en las plantas y se replican en células susceptibles. La molécula puede ser líneal (dando lugar a una estructura en horquilla) o circular (de forma aplanada); en todo caso, los pares de bases complementarios se unen dando como resultado la estructura de doble cadena, semejante a la del ADN. El primer viroide caracterizado, conocido como PSTVd, es una molécula de ARN de cadena simple que contiene 359 nucleótidos.
ma:2011-0054 #20 Otros biones acelulares Un virusoide o ARN satélite es una pequeña molécula patógena de ARN (ácido ribonucleico) que se encuentra encapsulada en ciertos virus de plantas con secuencias de ARN de mayor tamaño, lo que se denomina un virus asistente, el cual hospeda al virusoide y es necesario para que éste produzca una infección exitosa. El ARN circular del virusoide es extremadamente pequeño, solo unos cientos de nucleótidos, pues no necesita genes para nada salvo para replicarse. Los virusoides son similares a los viroides en tamaño y estructura, pero se diferencia en que los viroides no necesitan a un virus asistente. Aunque son estudiados en virología, no se les considera verdaderos virus sino partículas subvirales, por necesitar de otros virus para propagarse. Por esto, en las clasificaciones de virus aparecen con el término de "ARN satélite circular". De hecho, el término virusoide ha sido también usado genéricamente para referirse a todos los ácidos nucleicos satélites. Debido a que carecen de estructuras que le faciliten el ingreso a la célula que atacan, para ingresar dependen de un virus, por esto a los virusoides también se les denomina Satélites. Dentro de la célula, el virusoide se adueña de parte del mecanismo de síntesis de la bacteria, y la bacteria comienza a trabajar para el virusoide, formando numerosas réplicas de él, que quedan envueltas junto al material genético de los descendientes del virus que la célula infectada también esta formando. Así, el virus recién formado lleva en su interior una fracción de ARN que no le pertenece, al Satélite, o sea, al virusoide
mat:2011-0054 #21 Priones 1 Son agentes patógenos formados por una proteína ( proteína del prión o PPr ). Producen entre otras, la enfermedad de las "vacas locas" o encefalopatía bovina espongiforme. Esta proteína se acumula en el cerebro de animales enfermos , dando lugar a la estructura esponjosa de la corteza cerebral que da nombre a la enfermedad. Los priones, o las enfermedades producidas por priones, tienen un comportamiento sorprendente, por un lado se transmiten verticalmente , como cualquier enfermedad hereditaria típica, mientras que por otro lado se comportan de manera infectiva, transmitiéndose horizontalmente, mediante contagios que pueden darse entre individuos de distintas especies. La proteína del prión (PrP) normal, tiene una secuencia de aminoácidos, (estructura primaria) idéntica a la proteína del prión patógena. La diferencia entre las dos recae en la estructuras secundaria y terciaria. A pesar de que actualmente hay muy poca información sobre la función que tienen las proteínas Prpc (estado no patógeno), algunos experimentos han demostrado que tienen un papel activo en el correcto desarrollo neuronal, que es una proteína capaz de unir específicamente Cu2+ (procesos de oxidoreducción), y también se han relacionado los priones con proteínas de transducción de señales, la adhesión celular y la regulación y distribución de los receptores de acetilcolina.6
#22 Priones 2 Una vez obtenido el prion responsable de la patología, comenzó a dilucidarse su estructura. Se trata de una glicoproteína de 27-30 kD, que tiene la misma estructura primaria que una proteína similar presente en el cerebro de la oveja. La modificación estructural, no genética, se debe a un proceso de postraducción. Esto se presenta en funciones muy especiales. El gen codificando del prion Prpc (gen PRNP) se localiza en el brazo corto del cromosoma 20, no tiene intrones y es un gen autosómico dominante. Se expresa en el tejido neuronal, cardíaco, muscular, pancreático y hepático. El plegamiento erróneo de la Prpc a PrpSc confiere a la PrpSc dos propiedades que la diferencian de la Prpc: la resistencia parcial a la digestión por proteasas y su insolubilidad. Estas dos propiedades hacen que la PrpSc sea estable y la capacitan para poder formar agregados proteicos responsables de la acumulación de PrpSc en forma de placas amiloides en el tejido nervioso. Un prion es un tipo de proteína patógena (es decir, una molécula compleja que induce enfermedades), más específicamente una psialoproteína patógena, la cual tiene alterada su estructura secundaria, teniendo un incorrecto plegamiento de su estructura terciaria. A diferencia del resto de los agentes infecciosos (virus, bacterias, hongos etc...), que contienen ácidos nucleicos (ya sea ADN, el ARN, o ambos), un prion sólo esta compuesto por aminoácidos y no presenta material genético. El prion, palabra acuñada en 1982 por Stanley B. Prusiner al investigar una serie de enfermedades de carácter crónico e irreversibles que afectaban al sistema nervioso central, es un acrónimo inglés derivado de las palabras proteína e infección. Los priones son los responsables de las encefalopatías espongiformes transmisibles en una variedad de mamíferos, incluida la encefalopatía espongiforme bovina (EEB, también conocida como "enfermedad de las vacas locas") en el ganado y la enfermedad de Creutzfeldt-Jakob (ECJ) en humanos. Dichas proteínas mutadas forman agregados supramoleculares y son patógenas con plegamientos anómalos ricos en láminas beta, y autoreproducibles. Los priones se propagan mediante la transmisión de proteínas anómalas con mal plegamiento. Cuando un prion entra en un organismo sano, actúa sobre la forma normal del mismo tipo de proteína existente en el organismo, modificándola y convirtiéndola en prion. Estos priones recién formados pueden pasar a convertir más proteínas, provocando una reacción en cadena que produce grandes cantidades de la proteína prion. Todos los priones conocidos inducen la formación de amiloides plegado, en los que actúan polimerasas formando un agregado que consiste en apretadas hojas ß. El período de incubación de las enfermedades priónicas se determina por la tasa de crecimiento exponencial asociados con la replicación de priones, que es un equilibrio entre el crecimiento lineal y la rotura de los agregados (hay que tener en cuenta que la propagación del prion depende de la presencia de la proteína normalmente plegada en la que los priones pueden inducir plegamiento. Los organismos que no expresan la forma normal de la proteína priónica no pueden desarrollar o transmitir la enfermedad.
Antes de comenzar hablando de los virus conoceremos que es la
virología
La virología (rama de la microbiologia) es el estudio de los virus: su estructura, clasificación y evolución, su manera de infectar y aprovecharse de las células huésped para la reproducción del virus, su interacción con los organismos huéspedes, su inmunidad, la enfermedad que causan, las técnicas para su aislamiento,
Virus
Un VIRUS BIOLÓGICO se trata de una partícula infecciosa, un organismo que se replica a sí mismo en el interior de las células que ataca (células animales, vegetales o bacterianas).Podemos decir que no es un organismo totalmente vivo, ya que si analizamos la definición de ser vivo nos encontramos con que: nacen, crecen, se reproducen y mueren, de las cuales el virus por si solos solo nacen y mueren, necesitan una célula a la que infectar para crecer y evolucionar. No tienen metabolismo propio, necesitan una célula a la infectar, son “parásitos”.
ESTRUCTURA GENERAL DE LOS VIRUS
En los virus se distinguen las partes siguientes: 1.- Genoma vírico. Se compone de una o varias moléculas de ADN o de ARN, pero nunca los dos simultáneamente, Se trata de una sola cadena, abierta o circular, monocatenaria o bicatenaria. .
2.- Cápsida. Es la cubierta proteica que envuelve al genoma vírico. Está formada por proteínas globulares o capsómeros que se disponen de una manera regular y simétrica, lo que determina la existencia de varios tipos de cápsidas: icosaédricas, helicoidales y complejas. La función de la cápsida es proteger el genoma vírico y, en los virus carentes de membrana, el reconocimiento de los receptores de membrana de las células a las que el virus parasita.
3.-Envoltura membranosa. Formada por una doble capa de lípidos que procede de las células parasitadas y por glucoproteínas incluidas en ella cuya síntesis está controlada por el genoma vírico. Las glucoproteínas sobresalen ligeramente de la envoltura y tienen como función el reconocimiento de la célula huesped y la inducción de la penetración del virus en ella mediante fagocitosis.
La simetría es la forma que adopta un virus en el espacio y está dado por la estructura de su nucleocápside. La simetría puede ser helicoidal, icosaédrica o compleja. Los virus con simetría helicoidal presentan una nucleocápside cilindrica que puede estar extendida como en el virus del mosaico del tabaco. Este es un virus desnudo. En otros casos la nucleocápside está arrollada sobre sí misma y recubierta por una envoltura, como en el virusInfluenza. Los virus con simetría icosaédrica son poliedros regulares con 20 caras triangulares, 30 aristas y 12 vértices. Son ejemplos de virus con simetría icosaédrica desnudos el virus de la poliomielitis o losadenovirus. Los herpesvirus y togavirus son virus con simetría icosaédrica y recubiertos por una envoltura. Se denominan virus de simetría compleja a aquellos que presentan una nucleocápside helicoidal o icosaédrica recubierta por una envoltura laxa. Ejemplos de éstos virus son los poxvirus (forma de ladrillo), los rhabdovirus (forma de bala) y algunos bacteriófagos que poseen una cabeza con simetría icosaédrica y una cola con simetría helicoidal.
TAMAÑO DE LOS VIRUS
Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus. Algunos filovirus tienen una longitud total de hasta 1400nm.
continuación: FAMILIA DE VIRUS Hay muchas variedades de familias y de las más importante citaremos:
Familia Parvoviridae
- Virus de pequeño tamaño: son los más pequeños que existen. - DNA mc lineal 3 géneros: Gº Dependovirus Tienen virus defectivos: virus que no tienen capacidad de multiplicarse por sí solos, sino que dependen de otros virus: los adenovirus y los virus herpes. No provoca infecciones. Tienen antígeno común de grupo. Gº Densovirus Afectan a insectos. Interés: lucha biológica contra plagas. En concreto, afectan a ortópteros, lepidópteros, dípteros. Multiplicación tipo II de Baltimore, intranuclear. Exhibe poca información genética: 3-4 genes. Tiene una cápside proteica, y carece de envoltura de lípidos o glúcidos: es un virus desnudo. Resistente al éter y al cloroformo. Muy resistente a altas temperaturas (60º C durante 1 h.) y distintos pH (de 3-9) es muy difícil hacer una desinfección de este género. Gº Parvovirus Tienen tropismo por células en división. Provoca enteritis, panleucopenias (leucemias muy graves), depresión del SI. Gatos: tropismo por el cerebelo. Provoca hipoplasia celebelar (en cachorros, incluso fetos). Perros: tropismo por el cerebelo y también por el miocardio. Es patógeno para animales jóvenes. Son virus pantotropos, pero tienen apetencia por los tejidos descritos y el SI: Rango amplio de hospedadores: perros, gatos, cerdos, gansos, visones.
Familia Papovaviridae
- ADN bc, 1 hebra circular. - 5-8 genes - Desnudos resistentes al éter y al cloroformo - Morfología esférica, simetría cúbica (icosaédrica) - Estables a distintos pH y a distintas temperaturas (termoestables) - Mult. Intranuclear, ciclo de multiplicación: tipo I de Baltimore - Carecen de Ag de grupo, pero tienen antígenos comunes de género. - Cultivos celulares: efecto lítico y citopático - Virus transformantes: producen hiperplasias, amontonamientos celulares. - Cuerpos de inclusión Cowdry A - Virus hemoaglutinantes - Transmisión: contacto directo entre individuos y vectores (ácaros, sifonápteros: pulgas) - Tienen una elevada especificidad por el hospedador
Familia Adenoviridae
“Adeno” significa glándula. - ADN bc lineal. - 30 genes - Morf. esférica simetría cúbica. - Pequeño tamaño. - Tiene espículas, y también hemoaglutininas. - Multiplicación tipo I de Baltimore. - Tiene proteínas en la cápside, pero no envoltura (ni glúcidos ni lípidos) virus desnudo. - Resistente al éter y al cloroformo. - 252 capsómeros. - No tienen antígeno común de grupo pero sí antígenos comunes de género (interesante para vacunas). - Estables a distintas temperaturas y pH. - Efecto citopático, citolítico y también despegamiento celular en las células en cultivo. - Inducen hiperplasias.
Familia Herpesviridae
Incluidas dentro de la familia hay 3 subfamilias, con distintos géneros. En total hay más de 80 virus que afectan a todas las especies animales: aves, reptiles, anfibios, cerdos, caballos, rumiantes y animales salvajes. También al hombre. “Herpes” significa reptante. De todos los animales, el menos receptivo a los virus herpes son las ovejas. - ADN bc lineal - 160 genes - Morfología circular, simetría cúbica. - 162 capsómeros - Envoltura lipídica y proyecciones - Sensibles al éter y al cloroformo. - Partes: ácido nucleico, proteínas (cápside), lípidos y glúcidos. - Sensibles a la temperatura y al pH (lábiles).
Comprenden géneros que afectan tanto al hombre como a los animales. Son los virus de las viruelas, los de tamaño más grande que se conocen (visibles al microscopio óptico). Son virus modélicos, que han servido para el estudio de oncogénesis. Actualmente sirven como vectores vacunales: se les inyecta el ADN exógeno (de virus o de bacteria) y se puede producir lo que sea a partir de ese ADN. - Morfología y simetría compleja (tienen “cuerpos laterales”, de los que se desconoce la función). El interior de la célula tiene forma oval, y el exterior de ladrillo. - 300 genes. - Exhiben proteínas y enzimas.
DIAGNOSTICO DE LAS VIROSIS Síntomas Los síntomas son muy variables y dependen del virus y del órgano afectado. Muchos virus, como muchas bacterias, producen fiebre y síntomas respiratorios (tos, estornudos) o intestinales (náuseas, vómito, diarrea). Las virosis, aun cuando no sean de peligro, a menudo provocan fiebre elevada en los niños de corta edad. Diagnóstico Algunas infecciones virales, como la gripe, el resfriado común, la varicela son fáciles de identificar por sus síntomas, sin que se necesiten análisis de laboratorio. Para otras, como la hepatitis viral, el sida y la mononucleosis infecciosa, se suele analizar una muestra de sangre en busca de anticuerpos específicos contra el virus. La presencia de estos anticuerpos confirma el diagnóstico. En ciertos casos, los virus pueden cultivarse en tejidos celulares o bien el virus se identifica por su ácido nucleico, valiéndose de una técnica que recibe el nombre de reacción en cadena de la polimerasa (RCP). Las pruebas como el RCP o el cultivo celular se emplean cuando las de anticuerpos no tienen la precisión suficiente o cuando es necesario cuantificar con mayor exactitud el virus invasor.
ETAPAS DE LA REPLICACION
Adhesión: que depende de la interacción de proteínas de superficie del virus con receptores celulares específicos. Por ejemplo el receptor CD4 del linfocito T y la glicoproteína gp120 del virus HIV, las hemaglutininas de los virus influenza y los receptores de ácido acetil-neuramínico en la superficie de las células epiteliales, etc Penetración y decapsidación: la penetración a través de la membrana plasmática de la célula puede efectuarse por los siguientes mecanismos: por pasaje directo, por fusión de la membrana celular con la envoltura viral, o por endocitosis o viropexis, mecanismo más frecuente, es un fenómeno similar a la fagocitosis, en el cual el virus es incluído en una vacuola fagocítica. Luego de la penetración se produce la decapsidación, es decir la liberación del ácido nucleico viral de la nucleocápside. Penetración de los Bacteriófagos Inyección del material genético viral: Después de la adsorción, se produce un cambio configuracional en las proteínas de la placa basal, alguna de las cuales tienen actividad enzimática y producen un poro en la membrana citoplasmática de la célula. La vaina del fago se contrae y el material genético viral ingresa en la célula, mientras que la envoltura proteica queda en el exterior. Para entrar en una célula, los fagos se acoplan a receptores específicos en la superficie de la bacteria, que pueden ser lipopolisacáridos, ácidos teicoicos, proteínas o incluso flagelos. Por ello, cada fago solo podrá infectar ciertas bacterias según sus receptores. Puesto que los fagos no son móviles, dependen de encuentros al azar con los receptores adecuados en solución para poder infectar una bacteria. Parece que los bacteriófagos presentan una especie de jeringa mediante la cual introducen su material genético en el interior de la célula. Tras el reconocimiento del receptor adecuado, la cola y cuello del fago se contraen, quedando así el fago acoplado a la superficie celular. El material genético puede ser ahora introducido a través de la membrana o bien simplemente depositado sobre la superficie. No se descarta que pueda haber fagos con otros métodos diferentes para introducir su material genético en la célula.
MADURACION Y SALIDA DE VIRUS DESNUDOS Y ENVUELTOS. El desnudamiento ocurre simultáneamente o después de la penetración y comprende la eliminación de todas o parte de las cubiertas proteicas que rodean al ácido nucleico viral, de manera de dejar expuesto dentro de la célula el genoma viral desnudo o asociado a alguna enzima para que pueda expresarse.
En algunos casos sólo se remueve parte de la cápside viral y en esas condiciones el genoma expresa sus funciones. Para otros virus complejos, como por ejemplo los Poxvirus, el desnudamiento parece ocurrir en dos etapas: en la primera hay remoción de ciertas cubiertas proteicas por enzimas celulares, parte del genoma es tanscripto en esas condiciones y luego, en una segunda etapa, y a través de una proteína codificada por el virus, se produce el desnudamiento total del genoma.
En general el desnudamiento es un proceso difícil de estudiar y establecer con precisión sus características por la rapidez con que ocurre y la dificultad en separarlo temporalmente de la penetración. Es por ello que se suele denominar internalización del virión al conjunto de ambos eventos, penetración y desnudamiento.
SALIDA DE UN VIRUS ENVUELTO. Vemos aquí como los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes.
No todas las partículas víricas liberadas son infecciosas. La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
Los virus envueltos (como el VIH) son liberados de la célula huésped por gemación. Durante este proceso, el virus adquiere su envoltura, que es una parte modificada de la membrana plasmática del huésped. Maduración: de produce el ensamble de los ácidos nucleicos neoformados con las proteínas de la cápside. Liberación: los nuevos viriones salen de la célula por lisis de la misma, como el virus polio; o bien por brotación a través de la membrana celular, como los paramixovirus o arenavirus; o de la membrana nuclear, como los herpesvirus o del retículo endoplásmico, como los togavirus. MECANISMO DE ACCION DE LOS RETROVIRALES
Los antivirales son un tipo de medicamento usado para el tratamiento de infecciones producidas por virus. Tal como los antibióticos (específicos para bacteria), existen antivirales específicos para distintos tipos de virus. No sin excepciones, son relativamente inocuos para el huésped, por lo que su aplicación es relativamente segura. Deben distinguirse de los viricidas, que son compuestos químicos que destruyen las partículas virales presentes en el medio ambiente. Su eficacia (y a veces su toxicidad) están relacionadas con la competencia inmunológica del paciente. Su actividad es restringida al actuar sobre un “órgano diana” muy específico. Desarrollo de Resistencias. La mayoría solo son eficaces durante la fase de Replicación viral y no sobre los virus Latentes. La utilización óptima de los antivirales precisa de la realización de un diagnóstico clínico correcto.
Las drogas antirretrovirales actúan sobre diferentes pasos necesarios para la replicación viral; hasta la fecha, las más extendidas son aquellas drogas que actúan sobre la transcriptasa inversa y las drogas que actúan sobre las proteasas; aunque ya existen fármacos capaces de inhibir otros sitios tales como los inhibidores de la fusión y los de la integrasa. Los Inhibidores de la Reversotranscriptasa (IRT) alteran la función de la enzima llamada reversotranscriptasa que es la que utiliza el VIH para cambiar su mensaje genético a una forma que pueda ser fácilmente insertada dentro del núcleo de la célula infectada. Es la encargada de hacer una copia en ADN del ARN viral de manera que éste pueda insertarse en el ADN celular, mediante la acción de las integrasas, y así producir proteínas del VIH necesarias para la auto reproducción viral. Los IRT nucleósidos y los nucleótidos necesitan ser fosforilados para luego interactuar con un sustrato en el sitio de unión a la enzima y, cuando es incorporado, entonces termina la elongación de la cadena de ADN; mientras que los IRT no nucleósidos no necesitan ninguna conversión metabólica y bloquean directamente la reacción con la reverso transcriptasa por una interacción específica con el sitio de unión “sin sustrato” de la reversotranscriptasa del VIH. Los Inhibidores de las Proteasas (IP) son fármacos con una potente actividad antiviral. La proteasa del VIH es requerida para el clivaje del polipéptido gag-pol en sus partes funcionales y la inhibición de este clivaje trae como consecuencia una disminución en la producción de viriones maduros. O sea, estos medicamentos se parecen a los trozos de la cadena de proteína que la proteasa corta normalmente. Al adherirse a la proteasa, los IP previenen que la proteasa corte las cadenas largas de proteínas y enzimas en trozos más cortos que el virus necesita para reproducirse en nuevas copias de sí mismo. Aunque las cadenas largas no hayan sido cortadas en los trocitos correctos, las nuevas copias del VIH aún se construyen y continúan empujando a través de la membrana de la célula infectada. Pero estas nuevas copias del VIH son “defectuosas”, ya que no han sido formadas completamente, por lo tanto no pueden continuar infectando otras células.(1,2,3,4) INHIBIDORES DE FUSION Los inhibidores de la entrada evitan el ingreso del VIH a las células sanas, se adhieren a las proteínas de superficie de las células T o a las proteínas de superficie del VIH. Para que el VIH es una y pueda entrar a las células T, las proteínas de la capa externa del VIH deben unirse a las proteínas de superficie de las células T. Los inhibidores de la entrada evitan que este proceso ocurra. Algunos inhibidores de la entrada se adhieren a las proteínas gp120 ó gp41 de la capa externa del VIH. Otros inhibidores utilizan la proteína CD4 ó los receptores CCR5 ó CXCR4 de la superficie de las células T. Si los inhibidores de la entrada logran adherirse eficientemente a las proteínas, éstas quedan bloqueadas y entonces el VIH no puede unirse a la superficie de las células T e ingresar a las mismas.
El término acelular significa literalmente "sin células". Se aplica principalmente a los virus, pero también a provirus y viroides, que son entidades biológicas no formadas por células, a diferencia de bacterias, compuestas por células procariotas, y de hongos, vegetales y animales, formados por células eucariotas.
LOS VIROIDES I y II
son agentes infecciosos que, al igual que los virus, tienen un ciclo extracelular que se caracteriza por la inactividad metabólica y unciclo intracelular en el que causan infección al huésped susceptible, pero que a diferencia de los virus, los viroides no poseen proteínas ni lípidos y están constituidos por una cadena cíclica corta de ARN, circular o con forma de varilla, (que no codifica proteínas).1 Es importante decir que tanto su forma intracelular como extracelular son las mismas (ARN desnudo), los mecanismos por los cuales éstos logran causar infección están relacionados con la autocatálisis de su material genético. Estas pequeñas moléculas de ARN circular de cadena sencilla formadas por unos pocos cientos de nucleótidos. Carecen de cápsida y su tamaño es una milésima parte del de los virus más pequeños. - Aparecen con mayor abundancia en el núcleo de las células infectadas. - Sólo han sido detectados en plantas, en las que producen una gran variedad de enfermedades y pueden ser transmitidos tanto horizontal como verticalmente. - No existen ninguna evidencia de que los viroides son traducidos a proteínas, ni se conoce cómo causan las enfermedades. Lo que sí se sabe es que son replicados por los enzimas del huésped. - Algunas semejanzas entre la secuencia de nucleótidos de los viroides y ciertos secuencias del genoma de las plantas no traducidas (intrones) sugieren que los viroides evolucionaron a partir de los intrones.
Los ARNs satélites son pequeñas moléculas de tamaño similar al de los viroides de plantas (330-400 bases), que son empaquetados en cápsidas de determinadas cepas de virus (con cuyos genomios no muestran homologías). Se replican sólo en presencia del virus colaborador específico, modificando (aumentando o disminuyendo) los efectos patógenos de éste.
Los virusoides constituyen un grupo de ARNs satélites no infectivos, presentes en el interior de la cápsida de ciertos virus, con semejanzas estructurales con los viroides, replicándose exclusivamente junto a su virus colaborador.
LOS PRIONES I y II
Son entidades infectivas de un tipo totalmente nuevo y original, descubiertas por Stanley Prusiner en 1981, responsables de ciertas enfermedades degenerativas del sistema nervioso central de mamíferos (por ejemplo, el “scrapie” o prurito de ovejas y cabras), incluyendo los humanos (kuru, síndrome de Gerstmann-Straüssler, enfermedad de Creutzfeldt-Jakob). Se definen como pequeñas partículas proteicas infectivas que resisten la inactivación por agentes que modifican ácidos nucleicos, y que contienen como componente mayoritario (si no único) una isoforma anómala de una proteina celular. Tanto la versión celular normal (PrPC) como la patógena (PrPSc en el caso del “scrapie”) son glicoproteínas codificadas por el mismo gen cromosómico, teniendo la misma secuencia primaria. Se desconoce si las características distintivas de ambas isoformas estriban en diferencias entre los respectivos oligosacáridos que adquieren por procesamiento post-traduccional. A diferencia de los virus, los priones no contienen ácido nucleico y están codificados por un gen celular. Aunque se multiplican, los priones de nueva síntesis poseen moléculas de PrP que reflejan el gen del hospedador y no necesariamente la secuencia de la molécula del PrP que causó la infección previa. Se desconoce su mecanismo de multiplicación, y para discernir entre las diversas hipótesis propuestas quizá haya que dilucidar la función del producto normal y su posible conversión a la isoforma patógena infectiva. Recientemente se ha comprobado que, al menos algunas de las enfermedades por priones son simultáneamente infectivas y genéticas, una situación insólita en la Patología humana, habiéndose demostrado una relación entre un alelo dominante del PrP y la enfermedad de Creutzfeldt-Jakob. El gen del prión (Prn-p) está ligado genéticamente a un gen autosómico (Prn-i) que condiciona en parte los largos tiempos de incubación hasta el desarrollo del síndrome.
#1 Mat: 2011-0289 .. La Bacteriología es la rama de la Biología que estudia la morfología, ecología, genética y bioquímica de las bacterias así como otros muchos aspectos relacionados con ellas. Es de gran importancia para el hombre por sus implicaciones médicas, alimentarias y tecnológicas.
#2 Mat:2011-0289 -Origen de las celulas Eucariotas- El origen de los eucariontes es un complejo proceso que tiene un origen procariota. Si bien hay varias teorías que explican este proceso, según la mayoría de estudios se produjo por endosimbiosis entre varios organismos procariotas, en donde el ancestro principal protoeucariota es de tipo arqueano y las mitocondrias y cloroplastos son de origen bacteriano.
#3 Mat:2011-0289 -LA CELULA BACTERIANA- Las bacterias son microorganismos unicelulares que presentan un tamaño de unos pocos micrómetros (entre 0,5 y 5 μm, por lo general) y diversas formas incluyendo esferas (cocos), barras (bacilos) y hélices (espirilos). Las bacterias son procariotas y, por lo tanto, a diferencia de las células eucariotas (de animales, plantas, hongos, etc.), no tienen el núcleo definido ni presentan, en general, orgánulos membranosos internos. Generalmente poseen una pared celular compuesta de peptidoglicano. Muchas bacterias disponen de flagelos o de otros sistemas de desplazamiento y son móviles.
Las bacterias presentan una amplia variedad de tamaños y formas. La mayoría presentan un tamaño diez veces menor que el de las células eucariotas, es decir, entre 0,5 y 5 μm.
#4 Mat:2011-0289 -La pared gram positiva y gram negativa-
La gram positiva mide de unos 15 a 20 nanometros de diametro y la gram negativa mide 2 nanometros de diametro.
La pared gram positiva, poseen una pared celular interna y una pared de peptidoglucano(es un exoesqueleto que da consistencia y forma esencial para replicación y supervivencia de la bacteria), no tiene membrana externa, no tiene espacio periplasmático, la red de mureína está muy desarrollada y llega a tener hasta 40 capas, conservan el complejo yodocolorante.
La pared gram negativa, poseen una pared celular más compleja: pared celular interna, pared de peptidoglucano, bicapa lipídica externa, tiene membrana externa que forma un saco rígido alrededor de la bacteria, mantiene estructura y es barrera impermeable a macromoléculas, ofrece protección en condiciones adversas, tiene espacio periplasmático, pueden ser anaerobios o aerobios y poseen proteínas con concentraciones elevadas.
La tinción de Gram o coloración de Gram es un tipo de tinción diferencial empleado en Bacteriología para la visualización de bacterias, sobre todo en muestras clínicas. Debe su nombre al bacteriólogo danés Christian Gram, que desarrolló la técnica en 1884.
Se utiliza tanto para poder referirse a la morfología celular bacteriana como para poder realizar una primera aproximación a la diferenciación bacteriana, considerándose Bacteria Gram positiva a las bacterias que se visualizan de color moradas y Bacteria Gram negativa a las que se visualizan de color rosa o rojo o grosella.
Metodologia:
Recoger muestras. Hacer el extendido en espiral. Dejar secar a temperatura ambiente o fijarlas utilizando un mechero. Fijar la muestra con metanol durante un minuto o al calor (flameado 3 veces aprox.) Agregar azul violeta (cristal violeta o violeta de genciana) y esperar 1 minuto. Todas las células gram positivas se tiñen de color azul-púrpura. Enjuagar con agua. Agregar lugol y esperar entre 1 minuto. Enjuagar con agua. Agregar acetona y/o alcohol y esperar de 8 a 15 segundos aproximadamente (parte crítica de la coloración). Enjuagar con agua. Tinción de contraste agregando safranina o fucsina básica y esperar 45 segundos. Este tinte dejará de color rosado-rojizo las bacterias Gram negativas.
#6 Mat:2011-0289 -Ribosomas bacterianos-
Los ribosomas son las estructuras supramoleculares encargadas de la síntesis de proteínas, en un proceso conocido como traducción. La información necesaria para esa síntesis se encuentra en el ARN mensajero (ARNm), cuya secuencia de nucleótidos determina la secuencia de aminoácidos de la proteína; a su vez, la secuencia del ARNm proviene de la transcripción de un gen del ADN. El ARN de transferencia lleva los aminoácidos a los ribosomas donde se incorporan al polipéptido en crecimiento.
#7 Mat:2011-0289 -FORMAS Y AGRUPACIONES BACTERIANAS-
Las bacterias normalmente se multiplican por fisión transversal binaria. En muchas especies, las células hijas resultantes de un evento de división por fisión tienden a dispersarse por separado al medio, debido a la actuación de fuerzas físicas (movimiento browniano, cizallamiento, corrientes de convección, etc).
Los principales tipos de formas bacterianas son:
-Cocos (células más o menos esféricas)
-Bacilos (en forma de bastón, alargados),que a su vez pueden tener varios aspectos:cilíndricos,fusiformes y en forma de maza, etc.
Atendiendo a los tipos de extremos, éstos pueden ser:
-Redondeados (lo más frecuente)
-Cuadrados
-Biselados
-Afilados
-Espirilos: al igual que los bacilos, tienen un eje más largo que otro, pero dicho eje no es recto, sino que sigue una forma de espiral, con una o más de una vuelta de hélice.
*Vibrios: proyectada su imagen sobre el plano tienen forma de coma, pero en el espacio suelen corresponder a una forma espiral con menos de una vuelta de hélice.
Otros tipos de formas son filamentos, ramificados o no,anillos casi cerrados, y formas con prolongaciones (con prostecas).
#8 Mat:2011-0289 -AGRUPACIONES DE COCOS-
Algunos géneros bacterianos se agrupan de una manera característica. Esta agrupación se debe a la tendencia de las células hijas a permanecer parcialmente adheridas después de la división celular.
Los cocos pueden disponerse:
-De a pares y se los llama diplococos -Si se disponen en cadena se llaman estreptococos -Cuatro células esféricas conforman una tetrada -En forma de racimo o irrregular se llaman estafilococos -En paquetes cúbicos se denominan sarcinas
Espora en biología designa una célula reproductora generalmente haploide y unicelular. La reproducción por esporas permite al mismo tiempo la dispersión y la supervivencia por largo tiempo (dormancia) en condiciones adversas.
La espora produce un nuevo organismo al dividirse por mitosis sin fusión con otra célula, produciendo un gametofito pluricelular. La espora es un elemento importante en los ciclos vitales biológicos de plantas, hongos y algas. El término deriva del griego σπορά (sporá), "semilla". Las esporas se pueden clasificar según su función, estructura, origen del ciclo vital o por su movilidad.
El término también puede referirse a la etapa inactiva de algunas bacterias, lo que se denomina más correctamente endosporas y no son esporas en el sentido considerado aquí. La mayoría de los hongos producen esporas; aquellos que no lo hacen se denominan hongos asporógenos.
-Flagelos
Son filamentos proteicos responsables de la movilidad. Están constituidos por una proteína llamada flagelina. Se introducen en la membrana celular y se proyectan hacia fuera de la célula. Los flagelos se disponen de distinta manera alrededor del cuerpo bacteriano.
En algunas especies se encuentran en gran número y distribuídos sobre toda la superficie celular, a esta disposición se la llama peritrica. Cuando la bacteria tiene un único flagelo, en uno de los polos se denomina monotrica. Otras presentan un haz de flagelos en uno de los polos, se las llama lofotrica, las que presentan haces de flagelos en ambos polos, se las llama anfitricas, y las especies que carecen de flagelos se las denomina atricas. Los flagelos mueven a la célula mediante rotación y le permiten a la bacteria responder a nutrientes químicos o materiales tóxicos y acercarse o alejarse de ellos por el fenómeno llamado quimiotáxis.
Ciertas bacterias usan otros medios de locomoción, las espiroquetas se mueven pero no poseen flagelos, presentan un filamento axial, que les permite el movimiento. Otras como las Mixobacterias, lo hacen por deslizamiento.
I. Aeróbicas y Gram negativas, cocos y bacilos Familias 1. Pseudomonaceae son bacilos curvos o rectos, mótiles por flagelo polar, catalasa y oxidasa positiva. Contribuyen a degradar substancias quíímicas en el suelo, como por ejemplo pesticidas. Aquí encontramos el género Pseudomonas que se encuentra ampliamente distribuido en la tierra y en el agua. En este grupo incluimos la especie P. syringae que es un patógeno vegetal y causante de tumores a plantas. Xanthosomas todas las especies de este género son patógenas vegetales. 2. Rhizobiaceae Esta familia ocasiona hipertrofia en plantas, además de la formación de nódulos en raíces. Algunas especies pueden ocasionar tumores. Rhizobium forma nódulos en raíces de leguminosas y fija N2 atmosférico. Agrobacterium es importante en la ingeniería genética. Esta posee un plásmido con una región llamada T DNA que se inserta en el genoma de la planta huésped. 3. Methylococcaceae posee la habilidad para usar gas metano como fuente de carbono y energía bajo condiciones aeróbias o microaerofílicas. 4. Acetobacteraceae esta familia oxida etanol a ácido acético. Acetobacter y Gluconobacter son saprófitos que se encuentran en medios acídicos enriquecidos con azúcar o alcohol como flores, frutas, cervezas, vino, vinagre y otros. Poseen importancia industrial, ya queAzotobacter produce vinagre y Gluconobacter se utiliza para la manufactura de productos químicos. 5. Legionellaceae Legionella son oportunistas en humanos, pueden causar la muerte. Se transmiten por vía aérea 6. Neisseriaceae Neisseria son parásitos que habitan en las membranas mucosas de humanos y animales. N. gonorrhoeae y N. meningitidis son altamente patógenos, y este último causa meningitis cerebroespinal. Acetobacter son saprófitos que se encuentran en tierra, agua y basura. Son patógenos oportunistas que ocasionan una variedad de infecciones particularmente en pacientes hospitalizados. II. Anaeróbico facultativo, bacilos y gram negativo
-La glucosa-6-fosfato (glucosa 6P) interviene en las principales rutas del metabolismo de los glúcidos.
-El ácido oxalacético o su forma ionizada, el oxalacetato, es un importante metabolito intermediario de varias rutas metabólicas, entre ellas el ciclo de Krebs y la fotosíntesis C-4. Se puede sintetizar por dos reacciones anapleróticas al pasar el piruvato a oxalacetato por carboxilación o de aspartato a oxalacetato por transaminación.
-El ácido fosfoenolpirúvico es un ácido carboxílico. Su anión se denomina fosfoenolpiruvato y posee gran importancia en el metabolismo celulardebido a que posee un enlace fosfato de alta energía implicado en la glucólisis y gluconeogénesis.
-La enzima Alfa-cetoglutarato deshidrogenasa u oxoglutarato deshidrogenasa (succinil transferidora), EC 1.2.4.2, cataliza la siguientereacción utilizando como cofactor la tiamina difosfato.
#13 Mat:2011-0289 -FORMAS L-
-Formas L: Protoplastos y Esperoplastos Diferencias con los micoplasmas
Se denominan protoplastos las células bacterianas a las que se ha desprovisto totalmente de pared celular, mientras que esferoplastos son aquellas células bacterianas que poseen restos de pared.En el caso de Gram-positivas, la desorganización total de su pared, por lo que se obtienen protoplastos; en el caso de las Gram-negativas, quedan restos de membrana externa y de peptidoglucano atrapados en ella, por lo que se obtienen esferoplastos.
A diferencia de los micoplasmas son bacterias que carecen de pared celular. Debido a la ausencia de pared celular, los micoplasmas no son sensibles a los antibióticos que bloquean la síntesis de la pared celular, como la penicilina u otros antibióticos betalactámicos.
Las bacterias, como el resto de los seres vivos, necesitan una fuente de carbono para poder sobrevivir. El origen de esta fuente de carbono sirve como criterio de clasificación para las bacterias. Además, se necesita una fuente de energía que sirva para poder construir sus propias moléculas; el tipo de fuente de energía utilizada también sirve como criterio de clasificación.
Autótrofas: la fuente de carbono es inorgánica (CO2).
Fotolitotrofas: la energía utilizada es la luz. (Ejemplo: bacterias purpúreas del azufre).
Quimiolitotrofas: la energía utilizada es la liberada en reacciones químicas. (Ejemplo: bacterias incoloras del azufre).
Heterótrofas: la fuente de carbono es inorgánica.
Fotoorganotrofas: la energía utilizada es la luz. Quimioorganotrofas: la energía utilizada es la liberada en reacciones químicas. A este grupo pertenecen la mayoría de las bacterias.
Bacterias aerobias: son aquellas que necesitan oxígeno para su metabolismo. Realizan la oxidación de la materia orgánica en presencia de oxígeno molecular, es decir, realizan la respiración celular.
Bacterias anaerobias: son aquellas que no utilizan oxígeno molecular en su actividad biológica. La obtención de energía la realizan mediante catabolismo fermentativo. Se pueden distinguir dos grupos dentro de ellas:
Bacterias anaerobias facultativas: pueden vivir en ambientes con oxígeno o sin él.
Bacterias anaerobias estrictas: sólo pueden sobrevivir en ambientes carentes de oxígeno. Como ejemplo, Clostridium, causante del tétanos.
#17 Mat:2011-0289 -Mecanismo de resistencia bacteriana-
La mayoría de bacterias que hacen resistencia a algún antibiótico tienen la capacidad de eludir o evitar la acción del medicamento, gracias a 5 mecanismos:
-Producción de enzimas que destruyen fármaco activo: Este mecanismo es utilizado principalmente por los Staphylococos resistentes a la penicilina G, gracias a la producción de B lactamasas que destruyen el fármaco. -Cambio de permeabilidad al fármaco: Algunas bacterias como los estreptococos poseen una barrera natural de permeabilidad a los aminoglucósidos, que puede ser superada al administrar simultáneamente un fármaco activo contra la pared celular como por ejemplo la penicilina. -Alteran estructuralmente el "blanco del fármaco" , sucede principalmente con antibióticos como penicilinas y cefalosporinas, en las que la resistencia es ocasionada por alteración o perdida de la función de las PBP (Proteínas de unión a penicilinas). -Desarrollo de via metabólica diferente que pasa por alto la reacción inhibida por el fármaco: Hay bacterias que son resistentes a las sulfonamidas y no requieren PABA extracelular y, pueden utilizar Ácido fólico preformado -Desarrollo de enzima diferente que ejecuta función metabólica pero es menos afectada por el fármaco. Un ejemplo claro de esto se presenta en las bacterias resistentes al trimetropim, la ácido dihidrofolico reductasa se inhibe con mucha menor eficiencia que en bacterias susceptibles al trimetropim.
#1 Mat:2011-0289 -Mecanismo de Acción de Los Antibióticos-
Debido a que los antibióticos tienen efectos sobre una diversidad de bacterias, sus mecanismos de acción difieren basados en las características vitales de cada organismo y que, por lo general, son objetivos que no existen en las células de mamíferos.
Los agentes antimicrobianos actúan por una serie de mecanismos, muy diferentes entre ellos y cuyos blancos se encuentran en diferentes regiones de la célula atacada. Las diversas regiones de ataque antibacteriano en general son consideradas:
* Pared bacteriana * Membrana bacteriana * Síntesis de proteínas * Síntesis de ácidos nucleicos
La acción del agente antibacteriano es lograda mediante los siguientes mecanismos de acción:
* inhibición de la síntesis de la pared celular * inhibición de la síntesis de proteínas * inhibición del metabolismo bacteriano * inhibición de la actividad o síntesis del ácido nucleico * alteraciones en la permeabilidad de la membrana celular
Con cualquiera de estas acciones o con una combinación de ellas, el germen es incapaz de sobrevivir.
Las penicilinas son antibióticos del grupo de los betalactámicos empleados profusamente en el tratamiento de infecciones provocadas por bacterias sensibles. La mayoría de las penicilinas son derivados del ácido 6-aminopenicilánico, difiriendo entre sí según la sustitución en la cadena lateral de su grupo amino. La penicilina G o bencipenicilina fue el primer antibiótico empleado ampliamente en medicina; su descubrimiento ha sido atribuido a Alexander Fleming en 1928, quien junto con los científicos Ernst Boris Chain y Howard Walter Florey —que crearon un método para producir en masa el fármaco— obtuvo el Premio Nobel en Fisiología o Medicina en 1945.
Existe una gran diversidad de penicilinas. Algunas especies de hongos del género Penicillium sintetizan de forma natural penicilinas, como el primer tipo aislado, la penicilina G. No obstante, debido a la aparición de resistencias, se han desarrollado otras familias siguiendo básicamente dos estrategias: la adición de precursores para la cadena lateral en el medio de cultivo del hongo productor, lo que se traduce en la producción de penicilinas biosintéticas; y la modificación química de la penicilina obtenida por la fermentación biotecnológica, lo que da lugar a las penicilinas semisintéticas.
La penicilina natural o penicilina G es cristalina, totalmente soluble en agua, soluciones salinas y dextrosas isotónicas. El radical R, es el responsable de la sensibilidad a la hidrólisis por parte de las β-lactamasas, del enlace a proteínas transportadoras y del vínculo con las proteínas bacterianas PBP que transportan a la penicilina dentro de la célula.37 Además, se le asocia a la penicilina un dipéptido cisteína-valina, haciendo que la penicilina tenga la ideal afinidad por la enzima bacteriana transpeptidasa, la cual no se encuentra en el cuerpo humano y que permite la síntesis del peptidoglucano.
#3 Mat:2011-0289 Las cefalosporinas, son una clase de los antibióticos beta-lactámicos. Junto con las cefamicinas pertenecen a un subgrupo llamado los cefamos. Las cefalosporinas son similares a las penicilinas, pero más estables ante muchas β-lactamasas bacterianas y, por lo tanto, tienen un espectro de actividad más amplio.
as cefalosporinas son agrupadas en grupos llamados "generaciones" por sus características antimicrobianas. Las primeras cefalosporinas fueron agrupadas en la "primera generación" mientras que más adelante, cefalosporinas de espectro extendido fueron clasificadas como cefalosporinas de segunda generación. Cada nueva generación de cefalosporinas tiene más potencia frente a bacterias gram-negativas, características antimicrobianas perceptiblemente mayores que la generación precedente; actualmente se diferencian cuatro generaciones de cefalosporinas. Cabe destacar que las cefalosporinas de primera generación tienen mayor espectro de acción ante estafilococo y estreptococo que las generaciones más recientes.
Se clafisican:
Cefalosporinas de primera generación
Las cefalosporinas de primera generación tienen actividad predominante contra cocos grampositivos —Streptococcus y Staphylococcus—. Las cefalosporinas usuales no son activas contra cepas de Staphylococcus aureus resistentes a meticilina (SARM).
Cefalosporinas de segunda generación
Desde esta generación se amplia el espectro incluyendo anaerobios y microorganismos gram negativos. Las bacterias del género Klebsiella suelen ser sensibles
Cefalosporinas de tercera generación
Cefdinir Cefpodoxima Cefditoren pivoxilo Cefixima
Cefalosporinas de cuarta generación
Cefalosporinas de la cuarta generación tienen un mayor espectro de la actividad contra organismos gram-positivos que las cefalosporinas de la tercera generación. También tienen una mayor resistencia a beta-lactamasas que las cefalosporinas de la tercera generación.
Ceftobiprol : mayor actividad contra Staphylococccus aureus resistentes a meticilina, Streptococcus pneumonie resistente a penicilinas, Pseudomona aeruginosa y enterococos.
Ceftarolina : activa tanto frente a Staphylococcus aureus como estafilococos coagulasa negativos sensible y resistente a meticilina (MRSA) así como frente a las recientes cepas resistentes a vancomicina (VRSA) y resistentes a daptomicina
#4 Mat:2011-0289 Tetraciclinas, Amino glúcidos, Imidazoles, Macrolidos, Lincosamidas y Sulfamidas
La tetraciclina: es usada para tratar las infecciones provocadas por bacterias, incluyendo la neumonía y otras infecciones en las vías respiratorias; el acné; infecciones en la piel, los genitales y el sistema urinario; y la infección que causa úlceras estomacales (Helicobacter pylori). Este medicamento es también usado como una alternativa de otros medicamento para el tratamiento de la enfermedad de Lyme y para el tratamiento y prevención del carbunco (después de una exposición por inhalación). La tetraciclina pertenece a una clase de medicamentos llamados antibióticos de tetraciclina. Funciona al prevenir el crecimiento y diseminación de las bacterias. Los antibióticos no tienen ningún efecto en los resfríos, la gripe u otras infecciones víricas.
Los aminoglucósidos o aminósidos: son un grupo de antibióticos bactericidas que detienen el crecimiento bacteriano actuando sobre sus ribosomas y provocando la producción de proteínasanómalas. Actúan a nivel de ribosomas en la subunidad 30S bacteriana, y por ende, a nivel de síntesis de proteínas, creando porosidades en la membrana externa de la pared celular bacteriana. Tienen actividad especialmente en contra de bacterias Gram negativas y aeróbicas y actúan sinergísticamente en contra de organismos Gram positivos.
El imidazol: es un intermediario de la biosíntesis de la histidina que se forma desde el imidazol glicerol fosfato con la pérdida de agua. De estructura cristalina, su fórmula molecular es C3H4N2 y su masa molecular es 68.077 g/mol.
Pertenece a una familia de compuestos químicos aromáticos, caracterizada por disponer de un anillo pentagonal nitrogenado. Tiene propiedades anfotéricas, cualidad esta que mantendrán algunos de sus derivados.
Los macrólidos: son antibióticos naturales, semisintéticos y sintéticos que ocupan un lugar destacado en el tratamiento de infecciones causadas por bacterias intracelulares. Integran este grupo: eritromicina, claritromicina, azitromicina, espiramicina y roxitromicina.
Lincosamidas: Son antibióticos bacteriostáticos o bactericidas en dependencia de su concentración, el uso de este medicamento por v.i ha obtenido magníficos resultados en el tratamiento para bacterias anaerobias.
Una sulfamida : es una sustancia química sintética derivada de la sulfonamida, bacteriostática y de amplio espectro. Las sulfamidas se emplean como antibióticos,antiparasitarios y coccidiostáticos en el tratamiento de enfermedades infecciosas. Las sulfas son usadas tanto en medicina humana, como el sulfametoxazol, la sulfadiazina y la sulfabenzamida, así como veterinaria, como el sulfanitrato, el sulfapirazol y la sulfaquinoxalina. Las sulfonamidas fueron las primeras drogas eficaces empleadas para el tratamiento sistémico de infecciones bacterianas en el ser humano.
Las tetraciclinas constituyen un grupo de antibióticos, unos naturales y otros obtenidos por semisíntesis, que abarcan un amplio espectro en su actividad antimicrobiana. Químicamente son derivados de la naftacenocarboxamida policíclica, núcleo tetracíclico, de donde deriva el nombre del grupo.
Las tetraciclinas naturales se extraen de las bacterias del género Actinomyces. De Streptomyces aurofaciens se extraen la clortetraciclina y la demetilclortetraciclina, de Streptomyces rimosus se extrae la oxitetraciclina, y la tetraciclina, representante genérico del grupo, se puede extraer del Streptomyces viridifaciens, aunque también se puede obtener de forma semisintética. Una característica común al grupo es su carácter anfotérico, que le permite formar sales tanto con ácidos como con bases, utilizándose usualmente los clorhidratos solubles.
Actúan fundamentalmente como bacteriostáticos a las dosis habituales, aunque resultan bactericidas a altas dosis, generalmente tóxicas. Actúan por varios mecanismos:
Desacoplan la fosforilación oxidativa de las bacterias. Provocan una inhibición de la síntesis protéica en el ribosoma de la bacteria. Actúan inhibiendo la síntesis proteica al unirse a la subunidad 30 S del ribosoma y no permitir la unión del ácido ribonucléico de Transferencia (tRNA) a este, ni el transporte de aminoácidos hasta la subunidad 50 S.1 Existe también evidencia preliminar que sugiere que las tetraciclinas alteran la membrana citoplasmática de organismos susceptibles, permitiendo la salida de componentes intracelulares.5
Las resistencias bacterianas a las tetraciclinas son de aparición lenta, aunque mucho más rápida si se utiliza por vía tópica. El mecanismo bacteriano implicado puede ser mediante plásmido, lo que explica la reticencia a usar las tetracilinas en el ámbito hospitalario, para evitar la aparición de resistencias simultáneas a varios antibióticos. Existen resitencias cruzadas entre los miembros del grupo.
A. Viroides - Pequeñas moléculas de ARN circular de cadena sencilla formadas por unos pocos cientos de nucleótidos. Carecen de cáps ida y su tamaño es una milésima parte del de los virus más pequeños. - Aparecen con mayor abundancia en el núcleo de las células infectadas. - Sólo han sido detectados en plantas, en las que producen una gran variedad de enfermedades y pueden ser transmitidos tanto horizontal como verticalmente. - No existen ninguna evidencia de que los viroides son traducidos a proteínas, ni se conoce cómo causan las enfermedades. Lo que sí se sabe es que son replicados por los enzimas del huésped. - Algunas semejanzas entre la secuencia de nucleótidos de los viroides y ciertos secuencias del genoma de las plantas no traducidas (intrones) sugieren que los viroides evolucionaron a partir de los intrones. C. Priones - Las encefalopatías espongiformes transmisibles (TSEs) son enfermedades cerebrales degenerativas que ocurren en muchos mamíferos, incluido el hombre. En estas enfermedades, se desarrollan progresivamente cavidades que hacen que adquiera el aspecto de una esponja. El scrapie es una TSE que afecta a ovejas y cabras conocida desde hace 250 años. - En los años 80 aparecieron numerosos casos de vacas afectadas por una TSE en Gran Bretaña que pudieron relacionarse con la ingestión de piensos obtenidos a partir de ovejas afectadas por el scrapie. En los 90 apareció una variedad de la enfermedad que afectaba a algunas personas que habían consumido carne de vacas con TSE y que los medios de comunicación bautizaron con el nombre de “mal de las vacas locas”. - El kuru es otra TSE que afectaba a los nativos de una tribu de Nueva Guinea que practicaban canibalismo ritual. · Los priones son partículas infecciosas proteicas que provocan las encefalopatías espongiformes transmisibles. · Poseen la misma secuencia aminoacídica de una proteína normal, pero presentan una estructura diferente. · Inducen, por un mecanismo hasta ahora desconocido, la transformación de proteínas normales en anómalas. · Contradicen el dogma central de la biología molecular, según el cual el flujo de información es en todos los seres vivos: ADN à ARNm à Secuencia Aminoácidos à Estructura Tridimensional Proteínas. La teoría de los priones propuesta por Prusiner supone la existencia de dos plegamientos para una única secuencia de amino ácidos. Además, el replegamiento de la PrP normal por acción de la PrP patológica, implica un flujo de información de una proteína a otra a nivel de estructura terciaria.
Los ribosomas son complejos macromoleculares de proteínas y ácido ribonucleico (ARN) que se encuentran en el citoplasma, en las mitocondrias, en el retículo endoplasmatico y en los cloroplastos. Son un complejo molecular encargado de sintetizar proteínas a partir de la información genética que les llega del ADN transcrita en forma de ARN mensajero (ARNm). Sólo son visibles al microscopio electrónico, debido a su reducido tamaño (29 nm en células procariotas y 32 nm en eucariotas). Bajo el microscopio electrónico se observan como estructuras redondeadas, densas a los electrones. Bajo el microscopio óptico se observa que son los responsables de la basofilia que presentan algunas células. Están en todas las células (excepto en los espermatozoides). Los ribosomas no se definen como orgánulos, ya que no existen endomembranas en su estructura.[cita requerida]
En células eucariotas, los ribosomas se elaboran en el núcleo pero desempeñan su función de síntesis en el citosol. Están formados por ARN ribosómico (ARNr) y por proteínas. Estructuralmente, tienen dos subunidades. En las células, estas macromoléculas aparecen en diferentes estados de disociación. Cuando están completas, pueden estar aisladas o formando grupos (polisomas). Las proteínas sintetizadas por los ribosomas actúan principalmente en el citosol; también pueden aparecer asociados al retículo endoplasmático rugoso o a la membrana nuclear, y las proteínas que sintetizan son sobre todo para la exportación.
Tanto el ARNr como las subunidades de los ribosomas se suelen nombrar por su coeficiente de sedimentación en unidades Svedberg. En las células eucariotas, los ribosomas del citoplasma alcanzan 80 S. En plastos de eucariotas, así como en procariotas, son 70 S. Los ribosomas mitocondriales son de tamaño variado, entre 55 y 80 S.1
La Bacteriología es la rama de la Biología que estudia la morfología, ecología, genética y bioquímica de las bacterias así como otros muchos aspectos relacionados con ellas. Es de gran importancia para el hombre por sus implicaciones médicas, alimentarias y tecnológicas La bacteriología se clasifica en: • Bacteriología General • Bacteriología Agrícola • Bacteriología Alimenticia • Bacteriología Farmacéutica • Bacteriología Médica
Hay diversas clasificaciones para un mejor estudio de las bacterias: • Morfología • Organización celular • Presencia de núcleo • Tipo de nutrición • Tipo de reproducción • Tinción de Gram
La bacteriología se clasifica en: • Bacteriología General • Bacteriología Agrícola • Bacteriología Alimenticia • Bacteriología Farmacéutica • Bacteriología Médica
Hay diversas clasificaciones para un mejor estudio de las bacterias: • Morfología • Organización celular • Presencia de núcleo • Tipo de nutrición • Tipo de reproducción • Tinción de Gram
Hay diversas clasificaciones para un mejor estudio de las bacterias: • Morfología • Organización celular • Presencia de núcleo • Tipo de nutrición • Tipo de reproducción • Tinción de Gram
Cynthia Pamela de la Rosa --------------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3 DIAPOSITIVA: #1
QUE ES LA VIROLOGIA?
A traves de mis investigaciones pude encontrar la importancvia de la virologia, que es?, entre otros temas sobre la misma, a continuacion su definicion.
Virología es el estudio de virus y virus como agentes: clasificación y evolución, su estructura, sus formas de infectar y explotar las células para la reproducción del virus, las enfermedades que causan, las técnicas para aislar y la cultura de ellos y su uso en la investigación y terapia.
Virología es considerada una parte de la microbiología o de patología.
Una importante rama de la Virología es la clasificación de virus. Virus pueden clasificarse según la célula huésped que infectan: virus animales, virus de plantas, hongos virus y bacteriófagos (virus que infectan bacterias, que son los virus más complejos).
Otra clasificación utiliza la forma geométrica de su cápside (a menudo una hélice o un icosaedro) o la estructura del virus (por ejemplo, la presencia o la ausencia de un envolvente de lípidos). Gama de virus en tamaño de unos 30 nm a aproximadamente 450 nm, lo que significa que la mayoría de ellos no puede verse con microscopios de luces.
Cynthia Pamela de la Rosa ----------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3 DIAPOSITIVA: #2 ESTRUCTURAS GENERALES DE LOS VIRUS
Sabemos que un virus está compuesto de una molécula de ácido nucleico y una envoltura proteínica. Ésta es la estructura básica de un virus, aunque algunos de ellos pueden añadir a esto la presencia de alguna enzima, bien junto al ácido nucleico, como la transcriptasa inversa de los retrovirus, bien en la envoltura, para facilitar la apertura de una brecha en la membrana de la célula hospedadora.
A la unidad formada por el ácido nucleico y la envoltura proteínica se le denomina también virión.
El ácido nucleico es solamente de un tipo, ADN o ARN, nunca los dos.
Atendiendo al tipo de ácido nucleico se distinguen cuatro clases de virus:
- ADN de cadena doble - ADN de cadena sencilla - ARN de cadena doble - ARN de cadena sencilla
La envoltura proteínica recibe el nombre de cápsida. Está formada por unas subunidades idénticas denominadas capsómeros. Los capsómeros son proteínas globulares que en ocasiones tienen una parte glicídica unida. Se ensamblan entre sí dando a la cubierta una forma geométrica.
Cynthia Pamela de la Rosa ----------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3 DIAPOSITIVA: #3 SIMETRIA VIRALES
Atendiendo la forma de la cápsida, se pueden distinguir los siguientes tipos de virus:
- Cilíndricos o helicoidales: los capsómeros, que son de un solo tipo, se ajustan entorno una hélice simple de ácido nucleico. Un ejemplo lo constituye el virus del mosaico del tabaco.
- Icosaédricos: los capsómeros, que suelen ser de varios tipos, se ajustan formando un icosaedro regular (es decir, 20 caras triangulares y 12 vértices), y dejando un hueco central donde se sitúa el ácido nucleico fuertemente apelotonado. Algunos forman poliedros con más caras que el icosaedro, y algunos presentan fibras proteicas que sobresalen de la cápsida.
Un ejemplo lo constituyen los adenovirus, entre los que se encuentran los virus de los resfriados y faringitis.
- Complejos: con pequeñas variantes, responden a la siguiente estructura general:
- Una cabeza de estructura icosaédrica que alberga el ácido nucleico.
- Una cola de estructura helicoidal que constituye un cilindro hueco.
- Un collar de capsómeros entre la cabeza y la cola.
- Una placa basal, al final de la cola, con unos puntos de anclaje que sirven para fijar el virus a la membrana celular. De la placa salen también unas fibras proteicas que ayudan a la fijación del virus sobre la célula hospedadora.
Como ejemplo de este tipo de virus se encuentran la mayor parte de los virus bacteriófagos (que infectan las bacterias).
Cynthia Pamela de la Rosa ----------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3 DIAPOSITIVA: #4
Tamaño de los virus
Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus.
Su pequeño tamaño explica lo tardío del descubrimiento de estos agentes. La primera referencia sobre la existencia de los virus se debe al botánico ruso Dimitri Ivanovski en 1892. Este investigador buscaba el agente causante de la enfermedad denominada mosaico del tabaco, y llegó a la conclusión de que debía tratarse de una toxina o de un organismo más pequeño que las bacterias, pues el agente atravesaba los filtros que retenían las bacterias. Denominó a estos agentes patógenos virus filtrables.
~Picornaviridae... .virus ssRNA sentido-positivo..... coxsackievirus, virus de hepatitis A, poliovirus no-encapsulado
~Herpesviridae....dsDNA.....Virus de Epstein-Barr, herpes simple, tipo 1, herpes simplex, tipo 2, citomegalovirus humano, herpesvirus humano, tipo 8, Virus varicela-zoster.... encapsulado
~Hepadnaviridae dsDNA y ssDNA...... virus de hepatitis B encapsulado
~Flaviviridae...... virus ssRNA sentido-positivo..... virus de hepatitis encapsulado
~Retroviridae..... virus ssRNA sentido-positivo..... Virus de inmunodeficiencia humana (VIH) encapsulado
~Orthomyxoviridae...... virus ssRNA sentido-negativo..... virus de influenza encapsulado
~Paramyxoviridae..... virus ssRNA sentido-negativo..... virus del sarampión, virus de parotiditis, virus de parainfluenza, virus sincitial respiratorio encapsulado
Cynthia Pamela de la Rosa ----------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3 DIAPOSITIVA: #6
DIAGNOSTICO DE LA VIROSIS * METODOS DIRECTOS
Son aquellos que detectan:
1. El virus como agente infeccioso (aislamiento viral).
2. La presencia de antígenos virales (técnicas inmunológicas): Inmunofluorescencia (IF), Enzimoinmunoanálisis (EIA), Test de Aglutinación.
3.La presencia de ácidos nucleicos virales (PCR, etc.).
4.El virus como partícula viral (microscopía electrónica).
METODOS INDIRECTOS
Son aquellos que reconocen la respuesta inmune (humoral o celular) por parte del huésped: . Detección de anticuerpos específicos antivirales por técnicas inmunológicas (EIA, IFI, WB, etc.). Producción de anticuerpos in vitro.
En el curso de una infección varían las poblaciones de anticuerpos frente al agente infectante. En una primera fase la clase predominante suele ser IgM, mientras que con el transcurso del tiempo las IgM disminuyen hasta desaparecer o quedar a baja concentración residual y, en cambio, aumentan las IgG. La búsqueda de anticuerpos clase IgM es de utilidad para hacer diagnóstico de infección reciente en una sola muestra de suero extraída en el período agudo de la enfermedad. Este método se emplea para el diagnóstico de enfermedades como: Rubéola, Citomegalovirus, Hepatitis a virus A, etc.
Cynthia Pamela de la Rosa ----------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3 DIAPOSITIVA: #7
ETAPAS DE REPLICACION DEL VIRUS
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante.
Cynthia Pamela de la Rosa ----------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3 DIAPOSITIVA: #8
Penetracion de los Bacteriofagos
Los virus son moléculas de DNA o RNA rodeadas por una envoltura proteica que necesitan células viables para poder replicarse. Los virus utilizan la maquinaria metabólica de las células para sintetizar su material genético y proteínas de la envoltura. Existen distintos tipos de virus que pueden infectar células procariontes o células eucariontes. Los bacteriófagos o fagos son virus que se reproducen en células procariontes.
El genoma de los fagos puede ser RNA simple cadena (MS2, Qß), RNA doble cadena (phi 6), DNA simple cadena (phi X174, fd, M13) o DNA doble cadena (T3, T7, lambda , T5, Mu, T2, T4). Estos ácidos nucleicos pueden contener bases inusuales que son sintetizadas por proteínas del fago. En los T-pares el genoma no contiene citosina sino 5'- hidroximetilcitosina, mientras que en otros tipos de fago alguna de las bases esta parcialmente sustituida.
Cynthia Pamela de la Rosa ----------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3 DIAPOSITIVA: #9
Maduracion y salida de virus envueltos y desnudos.
Los virus cubiertos maduran por un proceso de gemación. Se insertan glucoproteínas de cubierta específica del virus en las membranas celulares y, a continuación, las nucleocápsides hacen gemación a través de la membrana a nivel de estos sitios modificados y, al hacerlo así, adquieren su cubierta. La gemación ocurre a menudo a nivel de la membrana plasmática y puede abarcar a otras membranas de la célula. Los virus cubiertos no son infecciosos hasta que han adquirido sus cubiertas. Por lo tanto, no se acumulan de manera típica viriones descendientes infecciosos en el interior de la célula infectada.
La maduración viral es a veces un proceso ineficaz. Pueden acumularse cantidades excesivas de componentes virales y participar en la formación de cuerpos de inclusión en la célula (ver antes). Como resultado de los profundos efectos dañinos de la replicación viral, se desarrollan por último efectos citopáticos celulares y la célula muere. Sin embargo, hay casos en que el virus no lesiona a la célula y se producen infecciones persistentes y prolongadas.os genomas virales recién sintetizados y los polipéptidos de la cápside se ensamblan para formar los virus descendientes. Las cápsides icosaédricas pueden condensarse en ausencia de ácido nucleico, en tanto que las nucleocápsides de los virus de simetría helicoidal no pueden formarse sin ARN viral.
No existen mecanismos especiales para la liberación de virus carentes de cubierta; las células infectadas acaban por experimentar lisis y liberar las partículas virales.
ESTEFANIA ROBLES LAJARA 2012-0358 MARTES DE 3-5 Virología esta es una parte de la microbiología y la patología que estudia los virus normal y también como agentes, su clasificación y evolución de los mismos, estructuras sus formas de infectar y explotar las células para la reproducción del virus, las enfermedades que causan, las técnicas para aislar y la cultura de ellos y su uso en la investigación y terapia Virus pueden clasificarse según la célula huésped que infectan: virus animales, virus de plantas, hongos virus y bacteriófagos (virus que infectan bacterias, que son los virus más complejos). Otra clasificación utiliza la forma geométrica de su cápside (a menudo una hélice o un icosaedro) o la estructura del virus (por ejemplo, la presencia o la ausencia de un envolvente de lípidos). Gama de virus en tamaño de unos 30 nm a aproximadamente 450 nm, lo que significa que la mayoría de ellos no puede verse con microscopios de luces. La forma y la estructura del virus ha sido estudiado por cristalografía de rayos X, microscopía electrónica y RMN. El sistema de clasificación más útil y más ampliamente utilizado distingue virus segun el tipo de ácido nucleico que utilizan como material genético y el método de replicación viral que emplean coaxial células huésped en la producción de más virus: • Virus de ADN (divididas en virus de ADN bicatenario y los virus de ADN monocatenario mucho menos común), • Virus ARN (dividieron en virus de ARN monocatenario de sentido positivo, virus de ARN monocatenario de sentido negativo y los virus de ARN bicatenario mucho menos común), • invertir transcribir virus (virus de ADN de transcripción inversa de bicatenario y monocatenario transcripción inversa RNA virus incluyendo retrovirus). También estudian '' partículas subvirales '', incluso más pequeñas que los virus infecciosas entidades: viroides (desnudas circulares moléculas de ARN infectando plantas), satélites (moléculas de ácido nucleico con o sin una cápside que requieren un virus auxiliar para la infección y reproducción) y priones (proteínas que pueden existir en una conformación patológica que induce a otras moléculas del prión para asumir esa misma conformación). Los taxones en Virología no son necesariamente monofiléticos. De hecho, las relaciones evolutivas de los distintos grupos de virus no están claras, y existen tres hipótesis sobre su origen: 1. Virus surgieron de materia no viviente, por separado y en paralelo a otras formas de vida, posiblemente en forma de autorreplicación RNA ribozimas similares a viroides. 2. Virus surgieron de la anteriores, más competentes celulares formas de vida que se convirtieron en parásitos a las células del huésped y posteriormente perdió la mayoría de sus funciones; ejemplos de tales diminutos parásitos procariotas son Mycoplasma y Nanoarchaeum. 3. Virus surgieron como partes del genoma de las células, probablemente transposones o plásmidos, que adquirieron la capacidad para "romper libre" desde el host de celda y infectan otras células. Por supuesto es posible que diferentes alternativas se aplican a grupos de virus diferente. De particular interés aquí es mimivirus, un virus gigante que infecta a las amebas y lleva gran parte de la maquinaria molecular tradicionalmente asociada con las bacterias. ¿Es una versión simplificada de una Célula procariota parasitaria o se originó como un simple virus que adquirido genes de su host? La evolución del virus, que a menudo se produce en concierto con la evolución de sus anfitriones, se estudia en el campo de la evolución viral. Mientras que el virus se reproduce y evolucionan, ellos no participan en el metabolismo y dependen de una célula huésped para la reproducción. La cuestión debatida a menudo de si están vivos o no es una cuestión de definición que no afectan la realidad biológica de los virus.
Los viroides al igual que los virus, tienen un ciclo extracelular que se caracteriza por la inactividad metabólica y unciclo intracelular en el que causan infección al huésped susceptible, pero que a diferencia de los virus, los viroides no poseen proteínas ni lípidos y están constituidos por una cadena cíclica corta de ARN, circular o con forma de varilla, (que no codifica proteínas) Los viroides son los agentes infecciosos de menor complejidad genética y estructural conocidos y representan una forma extrema de parasitismo. Están constituidos exclusivamente por moléculas de ARN de cadena simple, cerradas covalentemente o con forma de bastón de bajo peso molecular (246 a 1994 nucleótidos). Carecen de actividad de ARN mensajero y se replican de forma autónoma, utilizando el sistema de transcripción de la célula suceptible.2 Los viroides se encuentran, casi exclusivamente, en el núcleo de las células infectadas; se desconoce el modo en que se replican pero se sabe que el ARN que los constituyen no funciona como ARN mensajero y tampoco se traduce a enzimas que participen en su propia replicación. Los viroides no codifican para ninguna proteína y llevan a cabo su replicación en la célula hospedadora a través de la RNA polimerasa II codificada por la célula hospedadora aunque también se ha propuesto la posibilidad de que lo haga gracias a una RNA polimerasa dependiente de RNA presente en algunas células eucariotas (Cann, 2001). Dada su localización, se presume que causan la enfermedad interfiriendo con la regulación génica de la célula hospedadora en la etapa de corrección del ARN mensajero celular: particularmente en la eliminación de intrones y de empalmes de exones, presentando secuencias muy similares a los intrones que normalmente son cortados. Sus ARN se replican varias veces. Un prion1 es un tipo de proteína patógena (es decir, una molécula compleja que induce enfermedades), más específicamente una psialoproteína patógena, la cual tiene alterada su estructura secundaria, teniendo un incorrecto plegamiento de su estructura terciaria.2 A diferencia del resto de los agentes infecciosos (virus, bacterias, hongos etc...), que contienen ácidos nucleicos (ya sea ADN, el ARN, o ambos), un prion sólo esta compuesto por aminoácidos y no presenta material genético. A pesar de que actualmente hay muy poca información sobre la función que tienen las proteínas Prpc (estado no patógeno), algunos experimentos han demostrado que tienen un papel activo en el correcto desarrollo neuronal, que es una proteína capaz de unir específicamente Cu2+ (procesos de oxidoreducción), y también se han relacionado los priones con proteínas de transducción de señales, la adhesión celular y la regulación y distribución de los receptores de acetilcolina.6
La virología (rama de la microbiologia) es el estudio de los virus: su estructura, clasificación y evolución, su manera de infectar y aprovecharse de las células huésped para la reproducción del virus, su interacción con los organismos huéspedes, su inmunidad, la enfermedad que causan, las técnicas para su aislamiento, cultivo y su uso en investigación y terapia. La virología es considerada un sub-campo de la microbiología y la medicina. La virologia también es una rama taxonómica de la biología para estudiar los virus y todos sus componentes. Su investigación incluye:
• La replicación viral • Los patógenos virales. • La inmunología viral. • Las vacunas virales. • Los métodos de diagnóstico. • La quimioterapia antiviral. • Las medidas de control de una infección. • Los diferentes signos que manifiestan la presencia de virus.
El Ciclo De Replicación De Los Virus consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden sobrevivir por si mismos en el exterior (véase Virus). Para reproducirse se introducirán en células donde controlarán sus mecanismos reproductivos.
El origen de los eucariotas se encuentra en sucesivos procesos simbiogenéticos (procesos simbióticos que culminan en la unión de sus simbiontes, estableciéndose una nueva individualidad de los integrantes) entre diferentes bacterias. A principios del siglo XX, en 1909, el ruso Kostantin S. Mereschovky presentó la hipótesis según la cual el origen de los cloroplastos tendría su origen en procesos simbióticos.3 A parecidas conclusiones llegaron Kozo-Polyansky y Andrey Faminstyn (también de la escuela rusa) que consideraban la simbiogénesis “crucial para la generación de novedad biológica".4 En Francia, el biólogo Paul Portier, en 1918, y Ivan Wallin en Estados Unidos en 1927, llegaron a las mismas conclusiones. Trabajos que o bien pasaron inadvertidos (como los de la escuela rusa) o no fueron tenidos en cuenta (en el caso de Portier y Wallis) costando el prestigio profesional a sus proponentes. Lynn Margulis rescata estos trabajos y en 1967 en el artículo On origen of mitosing cells presenta la que llegaría a conocerse como Serial Endosymbiosis Theory (SET) (Teoría de la endosimbiosis seriada) en la que describe con concreción, mediante procesos simbiogenéticos, los pasos seguidos por las procariotas hasta la eclosión de las diferentes células eucariotas. Los tres pasos descritos por Margulis son:
Primera incorporación simbiogenética: Una bacteria consumidora de azufre, que utilizaba el azufre y el calor como fuente de energía (arquea fermentadora o termoacidófila), se habría fusionado con una bacteria nadadora (espiroqueta) habiendo pasado a formar un nuevo organismo y sumaría sus características iniciales de forma sinérgica (en la que el resultado de la incorporación de dos o más unidades adquiere mayor valor que la suma de sus componentes). El resultado sería el primer eucarionte (unicelular eucariota) y ancestro único de todos los pluricelulares. El núcleoplasma de la células de animales, plantas y hongos sería el resultado de la unión de estas dos bacterias. A las características iniciales de ambas células se le sumaría una nueva morfología más compleja con una nueva y llamativa resistencia al intercambio genético horizontal. El ADN quedaría confinado en un núcleo interno separado del resto de la célula por una membrana.5
Segunda incorporación simbiogenética: Este nuevo organismo todavía era anaeróbico, incapaz de metabolizar el oxígeno, ya que este gas suponía un veneno para él, por lo que viviría en medios donde este oxígeno, cada vez más presente, fuese escaso. En este punto, una nueva incorporación dotaría a este primigenio eucarionte de la capacidad para metabolizar oxígeno. Este nuevo endosombionte, originariamente bacteria respiradora de oxígeno de vida libre, se convertiría en las actuales mitocondrias y peroxisomas presentes en las células eucariotas de los pluricelulares, posibilitando su éxito en un medio rico en oxígeno como ha llegado a convertirse el planeta Tierra. Los animales y hongos somos el resultado de esta segunda incorporación.6
Tercera incorporación simbiogenética: Esta tercera incorporación originó el Reino vegetal, las recientemente adquiridas células respiradoras de oxígeno fagocitarían bacterias fotosintéticas y algunas de ellas, haciéndose resistentes, pasarían a formar parte del organismo, originando a su vez un nuevo organismo capaz de sintetizar la energía procedente del Sol. Estos nuevos pluricelulares, las plantas, con su éxito, contribuyeron y contribuyen al éxito de animales y hongos.7 El primer paso, al día de hoy, no se considera demostrado. A finales de los años ochenta y principio de los noventa diversos trabajos no admitían las homologías propuestas entre los flagelos de los eucariontes y de las espiroquetas.8 9 10 11 Margulis defiende que las asociaciones entre espiroquetas y protistas apoyan su teoría, y "la comparación de genes y genomas arqueobaterianos con secuencias de eucariontes han demostrado la relación filogenética de ambos grupos".
La Célula Bacteriana Las bacterias son células muy sencillas; carecen de núcleo y tampoco presentan orgánulos en el citoplasma. Se las denomina Procariotas. Son organismos unicelulares y se encuentran en todos los ecosistemas. Las bacterias son un numeroso grupo de seres vivos, con características muy diversas. En la clasificación de los Dominios, Woese, aparecen dos grupos de Procariotas, el Dominio Archaea, que engloba a los organismos más antiguos del Planeta, y el Dominio Bacteria, en el que se encuentran la gran mayoría de los organismos bacterianos actuales, también conocidos con el nombre de Eubacterias. Las formas que presentan las bacterias pueden ser:
Coco Bacilo Vibrión Espirilo
Las bacterias pueden presentarse como individuos sueltos, o formando colonias. Se pueden encontrar colonias de diplococos (bacterias redondeadas, de dos en dos), diplobacilos (bacterias alargadas, de dos en dos), estreptococos(cordones de bacterias redondeadas), estafilococos (masas laminares de bacterias redondeadas) o sarcinas(conglomerados tridimensonales de bacterias redondeadas).
La Pared Celular En Gram + Y Gram – BACTERIA GRAM POSITIVA
Tiene una capa gruesa de Peptidoglicano (mureina) y dos clases de ácidos teicoicos: Ácido Lipoteicoico que está en la superficie, empotrado en la capa de peptidoglicano y unido a la membrana citoplásmica. Y Ácido teicoico de la pared que está en la superficie y se une a la capa de peptidoglicano.
BACTERIA GRAM NEGATIVA
Tiene una capa delgada de Peptidoglicano (mureina) unida a una membrana exterior por lipoproteínas. La membrana exterior está hecha de proteína, fosfolípido y lipopolisacárido. En el lipopolisacárido, la porción de lípido está embebida en el fosfolípido y el antígeno O polisacárido entero se llama Endotoxina. La pared de la célula tiene poros llamados Porines para el transporte de substancias de peso molecular bajo.
Entre la membrana citoplásmica y la pared celular hay un espacio periplásmico con enzimas inactivadoras de antibióticos y proteínas de transporte.
Es un tipo de tinción diferencial empleado en microbiología para la visualización de bacterias, sobre todo en muestras clínicas. Debe su nombre al bacteriólogo danés Christian Gram, que desarrolló la técnica en 1884. Se utiliza tanto para poder referirse a la morfología celular bacteriana como para poder realizar una primera aproximación a la diferenciación bacteriana, considerándose Bacteria Gram positiva a las bacterias que se visualizan de color violeta y Bacteria Gram negativa a las que se visualizan de color rosa.
Son partículas constituidas por RNA y proteínas. El ribosoma bacteriano 70s consta de dos subunidades, pero es más pequeño que el de las células eucariotas. Es el lugar de síntesis de las proteínas, y el gran número de ribosomas refleja la importancia de esta función en la célula bacteriana. Se han encontrado algunas diferencias funcionales entre los ribosomas de las bacterias y los de las células eucariotas, tales diferencias han permitido encontrar compuestos con actividad antibiótica que actúan sobre el ribosoma bacteriano y no sobre el humano.
Formas Y Agrupaciones Bacterianas Los principales tipos de formas bacterianas son:
- cocos (células más o menos esféricas); - bacilos (en forma de bastón, alargados), que a su vez pueden tener varios aspectos:
- cilíndricos - fusiformes - en forma de maza, etc.
Atendiendo a los tipos de extremos, éstos pueden ser:
- redondeados (lo más frecuente) - cuadrados - biselados - afilados
espirilos: al igual que los bacilos, tienen un eje más largo que otro, pero dicho eje no es recto, sino que sigue una forma de espiral, con una o más de una vuelta de hélice.
vibrios: proyectada su imagen sobre el plano tienen forma de coma, pero en el espacio suelen corresponder a una forma espiral con menos de una vuelta de hélice. Otros tipos de formas filamentos, ramificados o no anillos casi cerrados formas con prolongaciones (con prostecas).
Estos distintos tipos de morfologías celulares deben de haberse originado por mecanismos evolutivos, a saber, por selección y estabilización adaptativa frente a las distintas presiones ambientales presentes en diferentes nichos ecológicos.
Son filamentos proteicos responsables de la movilidad. Están constituidos por una proteína llamada flagelina. Se introducen en la membrana celular y se proyectan hacia fuera de la célula. Los flagelos se disponen de distinta manera alrededor del cuerpo bacteriano. En algunas especies se encuentran en gran número y distribuídos sobre toda la superficie celular, a esta disposición se la llama peritrica. Cuando la bacteria tiene un único flagelo, en uno de los polos se denominamonotrica. Otras presentan un haz de flagelos en uno de los polos, se las llama lofotrica, las que presentan haces de flagelos en ambos polos, se las llama anfitricas, y las especies que carecen de flagelos se las denomina atricas. Los flagelos mueven a la célula mediante rotación y le permiten a la bacteria responder a nutrientes químicos o materiales tóxicos y acercarse o alejarse de ellos por el fenómeno llamado quimiotáxis.
Ciertas bacterias usan otros medios de locomoción, las espiroquetas se mueven pero no poseen flagelos, presentan un filamento axial, que les permite el movimiento. Otras como las Mixobacterias, lo hacen por deslizamiento.
Esporas
Algunas especies de bacterias Gram positiva, como las de los géneros Bacillus, Clostridium y Sporosarcina , disponen de un notable mecanismo adaptativo cuando se ven sometidas a privación de nutrientes en su medio ambiente: si los niveles de fuentes de C, N, o P caen por debajo de un umbral, esto constituye una señal a la célula de que se avecina un largo período de privación de nutrientes. Entonces, la célula se implica en una serie de complejos cambios genéticos, metabólicos, y estructurales, proceso de esporulación o esporogénesis, que conducen a la diferenciación, en el interior de la célula vegetativa original, de una célula latente, que incluye la región nuclear y algo de citoplasma, separado por una pared de estructura compleja, del resto de la célula madre, se forma así la endospora, que es capaz de permanecer en estado durmiente, varios decenios, incluso siglos. Las esporas son fácilmente diseminadas por el aire; cuando caen en medios ricos en nutrientes, se desencadena su germinación, se reinicia la actividad metabólica, de modo que cada espora genera una nueva célula vegetativa, capaz de dividirse y llevar a cabo una actividad metabólica normal. O sea, la esporulación se puede considerar como un potente proceso de supervivencia de ciertas bacterias Gram positivas cuando se enfrentan a condiciones severas de hambre de nutrientes. Las endosporas son formas de reposo (y no formas reproductivas), que representan una etapa del ciclo de vida de ciertas bacterias, y que se caracterizan por una estructura peculiar, diferenciada respecto de las células vegetativas, por un estado metabólico prácticamente detenido, y por una elevada resistencia a agentes agresivos ambientales, como desinfectantes químicos, radiaciones, el calor y la desecación. La posición de la bacteria puede ser central, terminal o subterminal; y puede ser del mismo tamaño que la célula bacteriana, mayor o menor. En el género Bacillus, la espora es de igual tamaño que la célula bactriana, en el género Clostridium es de mayor tamaño y deforma el cuerpo bacteriano.
Gránulos de reserva/inclusiones citoplasmáticas
Son acúmulos de sustancias orgánicas o inorgánicas que se originan dentro del citoplasma bajo determinadas condiciones de crecimiento. Su presencia y cantidad depende de la especie bacteriana, del nivel de actividad metabólica y de la composición del medio de cultivo. Los gránulos pueden ser de glucógeno, de lípidos, de volutita, que son reservorios de energía en forma de metafosfatos, de azufre, de proteínas, etc.
Las bacterias patógenas son una de las principales causas de las enfermedades y de la mortalidad humana, causando infecciones tales como el tétanos, la fiebre tifoidea, la difteria, la sífilis, el cólera, intoxicaciones alimentarias, la lepra y la tuberculosis. Hay casos en los que la etiología o causa de una enfermedad conocida se descubre solamente después de muchos años, como fue el caso de la úlcera péptica y Helicobacter pylori. Las enfermedades bacterianas son también importantes en la agricultura y en la ganadería, donde existen multitud de enfermedades como por ejemplo la mancha de la hoja, la plaga de fuego, laenfermedad de Johne, la mastitis, la salmonela y el carbunco.
Cada especie de patógeno tiene un espectro característico de interacciones con sus huéspedes humanos. Algunos organismos, tales como Staphylococcus o Streptococcus, pueden causar infecciones de la piel, pulmonía, meningitis e incluso sepsis, una respuesta inflamatoria sistémica que produce shock, vasodilatación masiva y muerte.[120] Sin embargo, estos organismos son también parte de la flora humana normal y se encuentran generalmente en la piel o en la nariz sin causar ninguna enfermedad.
Otros organismos causan invariablemente enfermedades en los seres humanos. Por ejemplo, el género Rickettsia, que son parásitos intracelulares obligados capaces de crecer y reproducirse solamente dentro de las células de otros organismos. Una especie de Rickettsia causa el tifus, mientras que otra ocasiona la fiebre de las Montañas Rocosas. Chlamydiae, otro filo de parásitos obligados intracelulares, contiene especies que causan neumonía, infecciones urinarias y pueden estar implicadas en enfermedades cardíacas coronarias.[121] Finalmente, ciertas especies tales como Pseudomonas aeruginosa, Burkholderia cenocepacia y Mycobacterium avium son patógenos oportunistas y causan enfermedades principalmente en las personas que sufren inmunosupresión o fibrosis quística.
Las infecciones bacterianas se pueden tratar con antibióticos, que se clasifican como bactericidas, si matan bacterias, o como bacterioestáticos, si solo detienen el crecimiento bacteriano. Existen muchos tipos de antibióticos y cada tipo inhibe un proceso que difiere en el patógeno con respecto al huésped. Ejemplos de antibióticos de toxicidad selectiva son el cloranfenicol y la puromicina, que inhiben el ribosoma bacteriano, pero no el ribosoma eucariota que es estructuralmente diferente.[124] Los antibióticos se utilizan para tratar enfermedades humanas y en la ganadería intensiva para promover el crecimiento animal. Esto último puede contribuir al rápido desarrollo de la resistencia antibiótica de las poblaciones bacterianas.[125] Las infecciones se pueden prevenir con medidas antisépticas tales como la esterilización de la piel antes de las inyecciones y con el cuidado apropiado de los catéteres. Los instrumentos quirúrgicos y dentales también son esterilizados para prevenir la contaminación e infección por bacterias. Los desinfectantes tales como la lejía se utilizan para matar bacterias u otros patógenos que se depositan sobre las superficies y así prevenir la contaminación y reducir el riesgo de infección.
FORMAS L Una Forma L es una variante bacteriana carente de pared celular (PC) o con PC defectuosa. Se estudia por primera vez en Streptobacillus moniliformis, donde se observa que produce en forma espontánea una serie de variantes capaces de reproducirse en forma de pequeños elementos filtrables carentes de PC o con PC defectuosa. Estos microorganismos reciben el nombre de Formas L (L: proviene de Lister Institute, Londres). Estas bacterias tienen una morfología colonial muy parecida a la de los Micoplasmas. Las Formas L se pueden presentar también en otras bacterias, siempre que la síntesis de PC esté alterada.
Formación La formación de las Formas L, es favorecida por acción de la Penicilina, por enzimas líticas que digieren el peptidoglicano o por concentraciones elevadas de sales como ocurre con la mayoría de las bacterias Grampositivas. Algunas bacterias Gramnegativas pueden convertirse a Formas L en medios con osmolaridad normal o fisiológica. Las Formas L son el equivalente morfológico de los Protoplastos y Esferoplastos, aunque el término de Formas L se limita a los organismos capaces de multiplicarse. Algunas Formas L, si se elimina el agente inductor, pueden revertir a la forma original (variante transitoria). Otras son Formas L estables. Durante muchos años se creyó que las Formas L eran Micoplasmas, pero se diferencian de ellos porque: a) Algunas Formas L pueden volver a poseer PC (formas inestables). Los Micoplasmas nunca poseen PC. b) Las membranas celulares de las Formas L carecen de esteroles y/o carotenoides, a diferencia de lo que ocurre con los Micoplasmas en los que estas sustancias brindan rigidez a su membrana celular. c) El análisis de homología de ácidos nucleicos demuestra que estos dos grupos no tienen ninguna vinculación taxonomica. La similitud de sus colonias y el parecido morfológico obedece a la ausencia de PC. d) Las Formas L de algunas especies bacterianas, requieren una concentración de sales en el medio que funcione como estabilizador osmótico, para mantener la integridad celular.
Protoplasto, del griego antiguo πρωτον (primero) + πλασσειν (moldear), inicialmente la palabra se refiere al primer cuerpo organizado de una especie.
En biología un protoplasto es una célula de planta, bacteria u hongo que ha perdido total o parcialmente su pared celular, para lo cual se usan mecanismos enzimáticos. En el caso de eliminar la pared con enzimas se obtienen: • Protoplastos - Cuando se elimina totalmente la pared celular. • Esferoplastos - Cuando sólo se elimina parcialmente la pared. Los protoplastos se obtienen a partir de bacterias Gram positiva y los esferoplastos de bacterias Gram negativa.
Esferoplasto
Un esferoplasto es una célula bacteriana desprovista de la mayor parte de su envoltura celular bacteriana mediante la adición de una penicilina. Los esferoplastos son muy frágiles y sensibles al estrés osmótico; tanto es así que se produce su lisis celular al transferirlos a un medio hipotónico. Los esferoplastos de algunas bacterias Gram negativas han sido empleados en investigación científica para dilucidar los mecanismos de acción de loscanales iónicos de su membrana biológica mediante la técnica del patch clamp, originalmente diseñada para estudiar la excitabilidad de células nerviosas como las neuronas. Para el análisis de los esferoplastos bacterianos, se crecen las células en presencia de inhibidores de la división celular, de modo que crezcan pero que no se dividan. Esto acentúa notablemente su tamaño y modifica su forma hasta estructuras bacilares. En este punto, se añade la penicilina, lo que provoca el debilitamiento de la pared y el colapso de la estructura hasta una forma esférica más estable (con menor proporción superficie/volumen), forma cómodamente analizable mediante patch-clamp. Es común que la bacteria sometida a este proceso haya sido modificada genéticamente añadiendo un gen para un canal iónico sobreexpresado, lo que aumenta su abundancia y facilita su análisis. Esta técnica ha sido empleada E. coli a fin de estudiar los canales mecanosensitivos (MscL, MscS, and MscM) desde 1987. El sistema se ha extendido para el análisis de canales heterólogos; los esferoplastos gigantes de E. coli son un modelo de expresión comparable al del oocito de anfibio Xenopus También es posible crear esferoplastos de levaduras como Saccharomyces cerevisiae mediante el empleo de enzimas como la zymolasa.
La nutrición es el proceso por el que los seres vivos toman el medio donde habitan las especies químicas que necesitanpara crecer. Según el tipo de nutrición las bacterias se clasifican en:
1- Autótrofas o Fotótrofas, son aquellas especies de bacterias coloreadas( verdes, pardas, rojas, purpúreas) que al poseer pigmentos fotosintétricos como la Bacterioclorofila, bacterioviridina, etc, son capáces de trasnformar sustancias inorgánicas simples como agua, fotones de luz solar, CO2 en alimentos orgánicos por Fotosíntesis. 2- Quimiosintéticas, son aquellas especies de bacterias que obtiene la energía química ( ATP ) a partir de la Oxidación de sustratos inorgánicos como agua, óxidos, hidróxidos, ácidos, sales minerales, entre ellas las mas conocidas son las bacterias fijadoras de N2 atmosférico como Nitrosomonas, Nitrobácter, otras como las bacterias del Azufre, Desulfovibrio sp que por quimiosíntesis obtienen el ATP a partir de la oxidación de compuestos azufrados, luego excretan una vaina azufrada que e ubica sobre la pared celular. 3- Heterótrofas, son aquellas bacterias que obtiene la energía química a partir de la incorporación o Absorción directa de alimentos sintetizados por otros seres vivos, el Heterotrofismo bacteriano puede ser:
a- De tipo Parásito, en la cuál las bacterias obtiene los nutrientes del organismo del cuál hospedan o parasitan. b- De tipo Saprófita, como la bacterias saprófitas que forman el eslabón de los Descomponedores en cadenas y redes tróficas, obtiene la energía química a partir de la descomposición de restos de organismos vegetales y animales en descomposición, entre ellas algunas especies mas conocidas están el Streptococus sanguis que por Saprofitismo de restos de alimentos que quedan entre los dientes origina la Placa bacteriana y los ácidos orgánicos son vertidos en el esmalte de los dientes causando las Caries dentales. c- De tipo Sombiótica, como la especie Eschrichia Coli que habita en el tracto intestinal y que obtiene los nutrientes en condiciones normales del sistema digestivo del hombre y de todos los mamíferos pero por compensación de los nutrientes tomados en forma directa provee o sintetiza al organismo "vitamina K" que actúa como anticoagulante evitando la coagulación de la sangre dentro de los vasos sanguíneos.
Es un proceso complejo, que puede darse aeróbica o anaeróbicamente dependiendo de la bacteria, se acompaña con la liberación de energía la cual servirá para la síntesis de compuestos. En las Bacterias la respiración puede ser de tipo:
En las bacterias Aerobias la respiración celular se realiza en la Cadena oxidativa o respiratoria asociada con la membrana plasmática que en ciertos casos se invagina formando una estructura llamada Mesosoma. A nivel de los Oxisomas localizados en el Mesosoma ya que carecen de mitocondrias los nutrientes son oxidados aeróbicamente liberando energía química y CO2.
En las Bacterias Anaerobias Facultativas (pueden o no fijar el O2 atmosférico) al respiración con O2 se realiza en los Oxisomas que forman las unidades fundamentales de la cadena oxidativa, utilizan la Glucólisis y luego la oxidación aerobia del ácido pirúvico en ATP y CO2, en tanto que aquellas bacterias que no fijan el O2 atmosférico la respiración es de tipo Anaerobia ( Fermentación) oxidando la Glucosa en condiciones anaerobias.
En las bacterias Anaerobias obligadas no poseen mecanismos para fijar el O2 atmosférico, por lo tanto la respiración en ellas es Anaerobia ( Fermentación).
Oxidan biológicamente la Glucosa por Glucólisis y luego trasnforman anaeróbicamente por Fermentación el ácido pirúvico en Etanal y posteriormente en Etanol (fermentación alcohólica).
Los testículos del toro y los ovarios de la vaca producen las células reproductoras por una serie de divisiones celulares que separan el número de cromosomas en una célula. El espermatozoide y el ovario contienen solamente un miembro del par de cromosomas.
Por lo tanto, las células de la vaca y del toro contienen 60 cromosomas (2n = 60), pero el espermatozoide en el semen y el óvulo en los ovarios contienen solamente 30 cromosomas (n=30, Figura 2). Los dos principios básicos de la transmisión de un rasgo (ejemplo sexo) son los siguientes:
1) Separación de los pares de cromosomas durante la formación de las células reproductoras;
2) Unión del espermatozoide con el óvulo para crear una nueva célula con un conjunto único de cromosomas.
Para 29 pares de cromosomas, ambos miembros son visualmente idénticos. De todas formas, para uno de los pares, un miembro es mucho más largo; es llamado cromosoma X, y el miembro más corto es llamado cromosoma Y. Todos los óvulos llevan el cromosoma X, pero el espermatozoide puede llevar ya sea el cromosoma X o el Y.
Rasgos cualitativos
Los rasgos cualitativos tienden a caer dentro de categorías discretas. Generalmente solo uno o unos pocos genes poseen un gran efecto sobre los rasgos cualitativos. El medio ambiente tene generalmente un pequeño papel al influenciar la categoría dentro de la que el animal cae. En este caso, el fenotipo de un animal refleja su genotipo. Ejemplos de rasgos cualitativos son:
* Color de pelo. * Defectos hereditarios como enanismo. * Presencia o ausencia de cuernos. * Tipo sanguíneo.
Rasgos cuantitativos
Los rasgos cuantitativos difieren de los cualitativos de dos formas importantes:
1) Se encuentran influenciados por muchos pares de genes. 2) La expresión fenotípica es influenciada más fuertemente por el medio ambiente que en el caso de los rasgos cualitativos.
El antibiograma es la prueba microbiológica que se realiza para determinar la susceptibilidad (sensibilidad o resistencia) de unabacteria a un grupo de antibióticos. Las técnicas de antibiograma son las utilizadas en el laboratorio de microbiología para estudiar la actividad de los antimicrobianos frente a los microorganismos responsables de las infecciones. Se considera como antimicrobiano cualquier sustancia con capacidad de matar o al menos de inhibir el crecimiento de los microorganismos y que sea susceptible de utilización como tratamiento en los pacientes. Pueden ser naturales, sintéticos o semisintéticos (modificación química de un compuesto natural). La historia moderna de los antibióticos comienza con el descubrimiento de sustancias presentes en unos microorganismos capaces de matar a otros microorganismos. La utilización de antibióticos supuso un avance enorme en la esperanza de vida de las personas que padecías procesos infecciosos, aunque desgraciadamente también supuso un aumento en los niveles de resistencia antibiótica.
El antibiograma tiene cuatro utilidades principales:
La utilidad básica del antibiograma es la instauración de un tratamiento antibiótico correcto al paciente. Es necesario conocer si el microorganismo responsable de la infección posee mecanismos que le confieran inmunidad frente a algún antibiótico para no incluirlo como terapia.
En cuanto al tratamiento el antibiograma no sólo es necesario en la instauración, también resulta útil en el seguimiento e incluso en la confirmación de tratamientos empíricos. En ocasiones la enfermedad infecciosa resulta grave y se comienza el tratamiento antes de conocer los datos de sensibilidad de la cepa. El antibiograma tiene que confirmar, o en su caso corregir el tratamiento. Otra aplicación de las técnicas de estudio de resistencia es la epidemiología. Es necesario detectar el aumento de los niveles de resistencia en los aislamientos clínicos para tomar medidas correctoras. Por otro lado también puede tener utilidad diagnóstica porque el perfil de resistencia puede en algún caso orientar en la identificación bacteriana.
Existen multitud de antimicrobianos en el mercado y el número sigue creciendo porque el desarrollo de más mecanismos de resistencia por parte de los microorganismos se traduce en un esfuerzo mayor por fabricar más y mejores antibióticos. No es necesario contrastar la eficacia de todos los antibióticos para cada aislamiento bacteriano. Los criterios que se siguen para seleccionar que antimicrobianos se ensayan respondes a factores de diversa índole: Factores microbiológicos: Tipo de agente infeccioso y mecanismos de resistencia descritos previamente en su especie. Factores farmacológicos: Tipo de antimicrobiano y parámetros de absorción, distribución y eliminación.
Factores del paciente: Tipo de infección. Factores de riesgo y estado general de salud. Situación inmunológica e hipersensibilidad. Experiencia anterior referente a los patrones de resistencia antibiótica más habituales para cada especie.
La resistencia bacteriana es un fenómeno creciente caracterizado por una refractariedad parcial o total de los microorganismos al efecto del antibiótico generado principalmente por el uso indiscriminado e irracional de éstos y no sólo por la presión evolutiva que se ejerce en el uso terapéutico.
El fenómeno de resistencia tiene un sustrato genético intrínseco o adquirido que se expresa fenotípicamente por mecanismos bioquímicos. De esta manera puede observarse la resistencia desde el ambiente biológico y otro el bioquímico.
Se conoce como resistencia natural a los mecanismos permanentes determinados genéticamente, no correlacionables con el incremento de dosis del antibiótico. Un ejemplo de esto es la resistencia de la Pseudomonas aeruginosa. a las bencilpenicilinas y al trimetoprin sulfametoxazol; bacilos gram negativos aeróbicos a clindamicina.
La resistencia adquirida aparece por cambios puntuales en el DNA (mutación) o por la adquisición de éste (plásmidos, trasposones, integrones).
En el primero se dan casos tales como la transformación de una Betalactamasa en una Betalactamasa de espectro extendido o como en el caso de mutaciones de los genes que codifican las porinas con el consecuente bloqueo del ingreso del antibiótico al interior del microorganismo.
Existen otras denominaciones de resistencia como son:
• Resistencia relativa o intermedia: ocurre un incremento gradual de la MIC (concentración inhibitoria mínima) a través del tiempo. Para obtener un efectoterapéutico es necesario alcanzar niveles séricos y tisulares adecuados. La susceptibilidad o resistencia del germen es en este caso dependiente de concentración.
• Resistencia absoluta: sucede un incremento súbito en la MIC de un cultivo durante o después de la terapia. Es inefectivo el incremento de la dosis clínica usual. Ejemplo de ello es la Pseudomonas spp. resistente a gentamicina y el Streptococcus pneumoniae altamente resistente a penicilina y uso de levofloxacina.
• Seudorresistencia: ocurre una resistencia in vitro pero una gran efectividad in vivo. Se denomina tolerancia antibiótica al fenómeno en el cual la diferencia entre la MBC (concentración bactericida mínima) y la MIC es muy grande lo cual ocurre con relaciones MBC/MIC mayores de 8 lo que permite la persistencia del microorganismo.
La virología (rama de la microbiologia) es el estudio de los virus: su estructura, clasificación y evolución, su manera de infectar y aprovecharse de las células huésped para la reproducción del virus, su interacción con los organismos huéspedes, su inmunidad, la enfermedad que causan, las técnicas para su aislamiento, cultivo y su uso en investigación y terapia. La virología es considerada un sub-campo de la microbiología y la medicina. La virologia también es una rama taxonómica de la biología para estudiar los virus y todos sus componentes.
Su investigación incluye:
• la replicación viral • los patógenos virales. • la inmunología viral. • las vacunas virales. • los métodos de diagnóstico. • la quimioterapia antiviral. • las medidas de control de una infección. • los diferentes signos que manifiestan la presencia de virus.
El Ciclo De Replicación De Los Virus consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden sobrevivir por si mismos en el exterior (véase Virus). Para reproducirse se introducirán en células donde controlarán sus mecanismos reproductivos.
La capsida, esta contiene una capa de proteína que la envuelve alrededor de un núcleo central de un producto químico altamente complejo llamado ácido nucleico, utilizado por el virus para su reproducción. Típicamente, la capsida se divide en subunidades llamadas los capsomeros. Las radiografias han mostrado que los virus tienen una capsida en forma de un sólido de 30 caras. EL Cuerpo, posee una simetría compleja, asociada a la capsida hay un vástago con una estructura que consiste en una vaina retráctil que rodea a un núcleo y es usada a modo de inyección. La cola, localizada al final del nucleo, es una placa espigada que lleva 6 fibras delgadas, que ayuden a asegurar al virus a sujetarse a la célula anfitrión, durante la invasión de la misma. En una serie de experimentos realizados en 1955,por Fraenkel Conrat y Williams ,demostraron que el virus del mosaico del tabaco (TMV),se formaba espontáneamente cuando se mezclaba una proteína purificada de la capa y su RNA geonómico ,al ser incubadas juntas, la estructura del TMV se regenera por si sola. A pesar de la gran variabilidad mostrada en las características del virus, todos se basan en algunas características básicas. Metodos de analisis La microscopia electrónica ha sido muy útil en dar información acerca de los virus con gran resolución, en dicho instrumento el nivel de la resolución es de 5nm (un nanometro 1nm equivale a 10^ -9 metros es decir 1 dividido 1000 millones de metro, para dar una cierta clase de perspectiva a esta dimensión diremos que: un átomo mide de 0,2-0,3 nm de diámetro una hélice de una proteína tiene 1nm de diámetro el ADN posee 2nm de diámetro La imagen obtenida con el microscopio electrónico puede darnos la forma de un virus con gran resolución, sin embargo para aumentarla a la escala atómica conveniente utilizar la cristalografía de rayos X, esto requiere que un virus pueda ser cristalizado, la cristalización de un virus fue descubierta en los años 30 y la estructura se puede determinar utilizando un modelo de difracción revelado por esta técnica , la primera estructura con resolución atómica de un virus fue realizada en 1978 y desde entonces muchas estructuras virales se han determinado. En algunos casos las estructuras se determinan con cristales de virus y en otros casos esta técnica no es posible. Un análisis bioquímico se puede utilizar para determinarse que componentes están presentes en el virus y en que relación. Los componentes individuales del virus, se cristalizan y sus estructuras se determinan. La clonación del gene y las técnicas de secuenciados facilitan el aislamiento de grandes cantidades proteínas purificadas y de los ácidos nucleicos para este propósito. La estructura total entonces es deducida construyendo el virus con estas subunidades, teniendo en cuenta también la información de otras técnicas tales como microscopia electrónica.
La simetría es la forma que adopta un virus en el espacio y está dad por la estructura de su nucleocápside. La simetría puede ser helicoidal, icosaédrica o compleja.
Los virus con simetría helicoidal presentan una nucleocápside cilindrica que puede estar extendida como en el virus del mosaico del tabaco. Este es un virus desnudo. En otros casos la nucleocápside está arrollada sobre sí misma y recubierta por una envoltura, como en el virusInfluenza.
Los virus con simetría icosaédrica son poliedros regulares con 20 caras triangulares, 30 aristas y 12 vértices. Son ejemplos de virus con simetría icosaédrica desnudos el virus de la poliomielitis o losadenovirus. Los herpesvirus y togavirus son virus con simetría icosaédrica y recubiertos por una envoltura.
Se denominan virus de simetría compleja a aquellos que presentan una nucleocápside helicoidal o icosaédrica recubierta por una envoltura laxa. Ejemplos de éstos virus son los poxvirus (forma de ladrillo), los rhabdovirus (forma de bala) y algunos bacteriófagos que poseen una cabeza con simetría icosaédrica y una cola con simetría helicoidal.
Es un taxón que afecta a vertebrados. Aden-(var.”adeno-“) Elemento prefijo del gr. “adén”, que significa “glándula”. Viriones isométricos no envueltos. Nucleocapside angular 70-90 nm diámetro. Simetría icosaedrica.
II- Virus de la peste porcina africana
Infecta a cerdos domesicos, facoceros, potamóquero de río y garrapatas. Causa fiebres hemorrágicas.
La peste africana se limitaba al continente que lleva su nombre hasta 1957 cuando la enfermedad se informó en otros países.
III- Arenaviridae
Virus que afecta a vertebrados.
Arena, viene del latín con la familia de términos del árido.
Estos virus son pequños, con envoltura y con forma icosaédrica.
V- Astroviridae
Virus que afecta a vertebrados.
Astro- es un prefijo lo cual significa estrellas.
Human astrovirus: Virones no envueltos. Nucleocapside isométrica. 27-30 nm diámetro. Simetría polihedrica. Con vértices.
VI- Baculoviridae
Virus que infecta vertebrados. Báculo: del latín, “palo”. Refiriéndose a la forma nucleocápside.
Son visibles en secciones de tejido infectado. Viriones ocluidos por cristales de proteínas o no ocluidos. Virion envuelto y pleomorfo forma de vara. Nucleocapsides en forma de vara. 200-450 nm long. Y 30-100 de diámetro.
VII- Badnavirus
Afecta a plantas
Etimología: Baciliforme Ba, DNA.
VIII- Barnaviridae
Infecta a hongos. Etimología: Baciliforme Ba, RNA. Sin envoltura, con un baciliformes T=1 simetría icosaédrica, aproximadamente 50 nm de longitud y 20 nm de gran tamaño.
IX- Birnaviridae
Afecta a vertebrados. Birna: para el ARN bipartito. No envueltos, de 60 nm de diámetro, virión icosaédricO con T=13 simetría.
X- Bromoviridae
Afecta a plantas. Bromo- porque infecta a Bromus inermis.
Viriones no envueltos, icosaédrico o baciliformes 26-35 nm de diámetro.
XI- Bunyaviridae
Afecta a vertebrados y a plantas. Viriones pleomorficos, esferoidales. “Bunyamwera” ciudad de Uganda donde se aisló.
Envueltos, esféricos. Diámetro de 80 a 120 nm.
XII- Caliciviridae
Virus que infecta a vertebrados y a invertebrados. “Calici” porque presenta depresiones calciformes.
Viriones no están envueltos y la capside es isométrica. Simetría icosaedrica.
XIII- Capillovirus
Infecta a plantas. Del latín “capella”, diminutivo de capa. El 3 es poliadenilado. Codifica por lo menos 3 proteínas.
Los virus son organismos tan pequeños, que su tamaño les permite pasar a través de los filtros que retienen bacterias. Las características básicas para identificar a un virus son: la composición del ácido nucleico, la disposición de la cápside, la presencia o ausencia de una envoltura, el tamaño del virus y el huésped.
DIAGNOSTICO VIRAL
1. Métodos para detectar virus. 1.1. Observación del efecto citopático, en cultivo de tejidos inoculados con ciertos virus. La etiología de un determinado síndrome viral puede demostrarse recuperando el agente causal en tejidos celulares de diversos tipos (por ej: riñón de mono, pulmón humano fetal, amnios humano o células tumorales humanas). Los huevos embrionados actualmente solo se utilizan para cultivar el virus cuando se desea preparar vacunas. El empleo de animales de experimentación es excepcional en el diagnóstico.
• Las células primarias de riñón de mono son excelentes para cultivar mixovirus, paramixovirus y muchos enterovirus; también para algunos adenovirus. • Las células diploides fetales humanas son células fibroblásticas en las que crece un amplio espectro de virus, como el de herpes simple, varicela zoster, adenovirus, picornavirus, y sólo en éstas pueden cultivarse los citomegalovirus. • Las células HEP-2 son epiteliales provenientes de un cancer humano, útiles para recuperar el virus respiratorio sincitial, adenovirus, y herpes simple. • Células mononucleares normales normales humanas de sangre periférica, que no suelen estar disponibles, permiten el cultivo del virus ( SIDA ) de la inmunodeficiencia, el coxsackie A y los togavirus. El crecimiento de un virus se detecta mediante la observación de cambios en la monocapa del cultivo celular, denominado efecto citopático (ECP). Cuando aparecen cambios en la morfología celular, lisis celular, vacuolización, formación de cincicios y existen cuerpos de inclusión, se produce un efecto citopático característico. Los cuerpos de inclusión son cambios histológicos que se producen en las células por los componentes virales o por variaciones en las estructuras celulares.
Continuación desarrollo del Tema: Diagnóstico Viral
Algunos virus no producen rápidamente ECP en las líneas celulares que se suelen utilizar en los laboratorios de virología clínica. No obstante se pueden determinar por otras técnicas, como la interferencia en la replicación de otros virus (por ej los picornavirus no pueden replicarse en células previamente infectados con el virus de la rubeola, lo que se denomina interferencia heteróloga).
1.2. Fijación de eritrocitos en cultivos celulares infectados con virus. En el caso de virus que dan lugar a este fenómeno, como mixovirus o virus de la parotiditis, se observa una hemaglutinación al adicionar eritrocitos a la monocapa de células. 1.3. Detección de antígenos virales. Cuando los virus se pueden detectar antes de desarrollar el efecto citopático, se utilizan anticuerpos monoclonales dirigidos contra antígenos tempranos y técnicas de inmunofluorescencia o inmunoperoxidasa. 1.4. Inmunofluorescencia directa, utilizando células del paciente, como leucocitos, orina o mucosa oral o genital, para buscar virus. 1.5. Detección de ácidos nucleicos virales por reacción de polimerasa en cadena, las cuales permiten amplificar una porción pequeña de ácido nucleico en un período de tiempo corto. 1.6. Microscopía electrónica. Cuando algunos virus comunes no se aislan con los métodos de cultivo habituales, ni pueden diagnosticarse por técnicas serológicas, pueden estudiarse con otras técnicas, como la microscopía electrónica. Tal es el caso de los calcivirus, los minirrotavirus, los astrovirus y los parvovirus del cerebro. Sin embargo, hay tres grandes dificultades para su uso: el instrumento es caro y es difícil su mantenimiento. Por otro lado, deben estar presentes grandes cantidades de particulas virales en la muestra para poder detectarse. Finalmente, la morfología no es suficientemente diferencial para un diagnóstico preciso.
2. Métodos para detectar anticuerpos específicos virales.
La mera presencia de anticuerpos específicos en contra de un determinado virus, no permite el diagnóstico de dicha infección. Debe tomarse en cuenta el cambio en los títulos en un lapso corto de tiempo, o ver si hay seroconversión. En muchas virosis la presencia de IgM sugiere infección reciente.
2.1. Fijación de complemento: Es una técnica estándar, pero de difícil ejecución. La muestra se procesa con el antígeno o el anticuerpo y el complemento; el complemento residual se determina mediante la lisis de eritrocitos cubiertos de anticuerpos que actúan como indicadores. Los anticuerpos medidos por este método se suelen desarrollar tardíamente en el desarrollo de la enfermedad. Los miembros de algunos grupos como adenovirus, influenza A y B, poseen antígenos comunes que se detectan por este método. Los demás grupos deben analizarse individualmente.
2.2. Neutralización: Cuando se incuba un virus con anticuerpos homólogos específicos de tipo, el virus no es capaz de producir la infección en un sistema de cultivos celulares indicadores. La respuesta de anticuerpos neutralizantes es específica del tipo devirus y se desarrolla al comienzo de los síntomas, elevándose los títulos rápidamente durando mucho tiempo. 2.3. Inhibición de la hemaglutinación: Se puede realizar con diversos virus que aglutinan selectivamente los eritrocitos de varias especies animales (por ej: pollo, cobaya y humanos). La capacidad de hemaglutinación del suero se inhibe por el suero humano específico. Los anticuerpos inhibidores de la hemaglutinación se desarrollan tras el comienzo de los síntomas, posteriormente se elevan y al final disminuyen. Esta prueba es útil para detectar virus de rubeola y para determinar el estado inmunitario.
1- El primer paso en la infección viral es la absorción a la membrana de la célula susceptible por medio de la adhesión de ligandos virales (proteínas de la capside o glucoproteínas de las espículas, por ejemplo a receptores superficiales de la célula). La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus. Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor. 2- Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre lasproteínas de la cápsida. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica.
Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras:
• Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma. • Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma.
Los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos:
• Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma. • Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
3- Según la duración de la fase de eclipse, se suelen distinguir dos modalidades de ciclo infeccioso de un virus: • Ciclo ordinario: el ácido nucleico vírico procede inmediatamente a la transcripción de su mensaje genético en los ARN necesarios para su multiplicación, y prosigue rápidamente el ciclo vital. Este tipo de ciclo es el más extendido en la naturaleza. • Ciclo lisogénico: fue descubierto por André Lwoff en bacteriófagos. El ADN vírico se cierra por sus extremos generando un ADN circular. Este ADN se inserta en el ADN bacteriano en un lugar específico en el que la secuencia de nucleótidos bacterianos es semejante alguna región del ADN vírico.
La bacteria prosigue sus funciones vitales sin que el virus realice ninguna acción, y cuando el ADN bacteriano se duplica también lo hace el ADN vírico, de manera que el genoma del virus pasa a las dos bacterias hijas. La multiplicación bacteriana puede seguir durante generaciones sin que el virus se manifieste. Pero ante una alteración de las condiciones ambientales, el ADN vírico se separa del bacteriano y prosigue entonces las restantes fases de ciclo infeccioso, produciendo la muerte de la bacteria y nuevos ejemplares del virus.
4- La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínasvíricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores. Losribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada.
Los antivirales son un tipo de medicamento usado para el tratamiento de infecciones producidas por virus. Tal como los antibióticos (específicos para bacteria), existen antivirales específicos para distintos tipos de virus. No sin excepciones, son relativamente inocuos para el huésped, por lo que su aplicación es relativamente segura. Deben distinguirse de los viricidas, que son compuestos químicos que destruyen las partículas virales presentes en el medio ambiente. Descripción Muchos de los antivirales disponibles actualmente están diseñados para ayudar el tratamiento del VIH (virus del sida), herpesvirus, productores de la varicela, el herpes labial, el herpes genital, etc. y los virus de la hepatitis B y C, que pueden causar cáncer de hígado. Los investigadores están trabajando actualmente para extender el rango de antivirales a otras familias de patógenos.
Características de un antiviral
Para que un fármaco antiviral sea considerado para el tratamiento de una infección viral debe cumplir ciertos requisitos indispensables. El fármaco debe ser específicamente activo contra el virus“target” (blanco), inhibiendo algún paso esencial de su metabolismo y debe poder debilitar a las cepas resistentes que puedan surgir. Además, debe tener ciertos parámetros que permitan al fármaco ser de alta biodisponibilidad oral, rápida penetración en los tejidos infectados y atóxicos para las células normales y el organismo en general a corto y largo plazo. El fármaco debe ser transformado a su forma activa solamente en células infectadas, debe ser excretado sin ser metabolizado o sin generar metabolitos tóxicos, y debe tener baja frecuencia de administración. Finalmente, el fármaco debe tener bajo costo para que pueda estar disponible para toda la población. Todas estas características, entre otras, dificultan el descubrimiento de nuevos fármacos efectivos para el tratamiento de las enfermedades virales.
Los fármacos antirretrovirales no matan al virus de la inmunodeficiencia humana (VIH). Ni siquiera logran dormirlo. Lo 'atontan' un poco y lo mantienen en un estado de reposo, pero que le permite, de vez en cuando, salir a 'pasear' por las células del organismo humano. Lo ha descubierto un equipo de investigadores españoles, que pone así fin a una de las controversias médicas más extendidas en el campo del sida.
Durante años, la teoría más apoyada ha sido la que consideraba que el cóctel de fármacos impedía la replicación del virus. No era capaz de eliminarlo, pero sí podía evitar que siguiera infectando. Otra hipótesis, más minoritaria, indicaba que a pesar de la terapia, el virus podía mantener algo de actividad. Y es esta última la que finalmente se ha confirmado, según la investigación que publica 'Nature Medicine' y que ha sido realizada por expertos del Instituto de Investigación del Sida IrsiCaixa, ubicado en el Hospital German Trias i Pujol de Badalona. Lo que han visto los investigadores, dirigidos por Javier Martínez Picado, es que algunos pacientes infectados por el VIH, a pesar de que presentan una carga viral indetectable gracias al tratamiento y que parecen tener la infección controlada, cuentan con una pequeña proporción del virus que sigue siendo infeccioso. Un descubrimiento que, según sus autores, "tiene importantes implicaciones clínicas, ya que podría explicar por qué, a pesar de que los pacientes permanecen en terapia durante muchos años, los fármacos no permiten curar la infección de manera definitiva". Martínez Picado lo aclara de forma sencilla a ELMUNDO.es: "Un paciente infectado sin tratamiento es como un bosque en llamas. Con los fármacos logramos apagar el fuego, pero quedan rescoldos y basta con un pequeño golpe de aire para que vuelva a incendiar. Esto es lo que hace el virus. Está apagado, pero no muerto". Carga viral indetectable Para llegar a estas conclusiones, el equipo analizó a 69 pacientes que seguían la terapia antirretroviral y que tenían carga viral indetectable (cantidad de virus en sangre) durante una media de cinco años. A 45 de ellos se les intensificó el tratamiento añadiendo otro fármaco, el Raltegravir, el primero de una nueva familia de medicamentos antisida, que bloquea la acción de la integrasa (una enzima que utiliza el VIH para copiar su material genético en el ADN de las células del paciente). Los otros participantes siguieron con su terapia antirretroviral normal.
VIROIDES Los viroides son agentes infecciosos que, al igual que los virus, tienen un ciclo extracelular que se caracteriza por la inactividad metabólica y unciclo intracelular en el que causan infección al huésped susceptible, pero que a diferencia de los virus, los viroides no poseen proteínas ni lípidos y están constituidos por una cadena cíclica corta de ARN, circular o con forma de varilla, (que no codifica proteínas).1 Es importante decir que tanto su forma intracelular como extracelular son las mismas (ARN desnudo), los mecanismos por los cuales éstos logran causar infección están relacionados con la autocatálisis de su material genético. En sí constituyen una etapa primitiva de los virus. El primer viroide fue descubierto por T. O. Diener en 1978, al intentar identificar el agente causal de una enfermedad que inicialmente supuso era inducida por un virus llamada "la enfermedad del tubérculo fusiforme de la patata" (potato spindle tuber o PSTVd) La nomenclatura de los viroides incluye el sufijo “Vd” para distinguirlos de los virus. El segundo viroide en ser descubierto fue el agente causal de la exocortis de los cítricos (citrus exocortis viroid, CEVd). Actualmente se conocen más de 200 viroides.
Satélites (Schneider): Virus asociados a otros virus de los que dependen para su multiplicación y capacidad para causar infección. Sólo de vegetales. Son incapaces de multiplicarse sin la concurrencia de un virus y controlan la capacidad de los virus. Actúan como parásitos de virus. 2- Virusoides o sateloides (Fransky): pequeñas hebras de ARN monocatenario circular, localizados dentro de otros virus ARN. 3- Elementos transponibles o transposones. 4- Plásmidos
Un prión es un agente infeccioso que se compone principalmente de proteínas. Hasta la fecha, todos los agentes que han sido descubiertos propagan por transmitir un Estado de mis-folded de proteínas; la proteína misma no máquinas y el proceso depende de la presencia del polipéptido en el organismo hospedador. La mis-folded forma de la proteína del prión ha sido implicada en una serie de enfermedades en una gran variedad de mamíferos, incluyendo encefalopatía espongiforme bovina (EEB, también conocida como "enfermedad de las vacas locas") en el ganado y la enfermedad Creutzfeldt - Jakob (ECJ) en los seres humanos. Todas las enfermedades priónicas conocidos afectan a la estructura del cerebro o de otros tejidos neuronales, y todos están actualmente intratable y siempre son fatales. En general uso de prión se refiere a la unidad teórica de la infección. En notación científica, PrPC se refiere a la forma endógena de la proteína del prión (PrP), que se encuentra en multitud de tejidos, mientras que PrPSc se refiere a la forma mal doblada de PrP, que es responsable de la formación de placas de amiloide y la neurodegeneración.
Priones son hipótesis a infectar y propagarse por células anormalmente en una estructura que es capaz de convertir las moléculas de la proteína normales en forma anormalmente estructurada. Todos conocido priones inducen la formación de un pliegue de amiloide, en los que la proteína polymerises en un agregado que consta de láminas beta estrechamente empaquetados. Esta estructura alterada es muy estable y se acumula en el tejido infectado, causando la muerte de células y daño del tejido. Esta estabilidad significa que los priones son resistentes a la desnaturalización por agentes químicos y físicos, dificultando la eliminación y contención de estas partículas. Mostrando el comportamiento del prión-tipo de proteínas también se encuentran en algunos hongos, que ha sido útil para ayudar a comprender los priones mamíferos. Priones hongos, sin embargo, no parecen causar enfermedades a sus anfitriones y incluso pueden conferir una ventaja evolutiva a través de una forma de herencia basados en proteína. La palabra prion es una palabra compuesta que se deriva de las letras iniciales de las palabras proteinaceous'' y menfectious'', con -en añadido por analogía a la palabra virien.
2011-0119 dios te bendiga Lista de arbovirus (incompleta)
Familia Togaviridae El virus chikungunya (CHIKV) (Vector: Mosquito) El virus O'nyong-nyong (Vector: Mosquito) El virus del Río Ross (Vector: Mosquito) El virus Sindbis (Vector: Mosquito)
Familia Flaviviridae[editar] Fiebre amarilla (Vector: Mosquito) Dengue (Vector: Mosquito) Encefalitis japonesa (Vector: Mosquito) Virus Kunjin (Vector: Mosquito) Enfermedad de Kyasanur (Vector: Garrapata) virus Usutu (USUV) (Vector: Mosquito) Virus del Nilo Occidental (Vector: Mosquito) Encefalitis de San Luis (Vector: Mosquito)
Familia Bunyaviridae La Fiebre del Valle del Rift (Vector: Mosquito) Encefalitis de La Crosse (Virus de La Cross) (Vector: Mosquito) Fiebre por Orepuche (Virus de Orepuche) (Vector: Mosquito) Virus Toscana (VTOS) (Fiebre Pappataci) (Vector: Mosquito) Virus Trivittatus (Vector: Mosquito)
Familia Reoviridae Lengua azul (Vector: Mosquito) Fiebre del Colorado por garrapatas (Vector: Garrapata) Virus Lipovnik (LIPV) (Vector: Garrapata) Virus Tribec (Vector: Garrapata) Virus Eyach (Vector: Garrapata)
Los fármacos antirretrovirales no matan al virus de la inmunodeficiencia humana (VIH). Ni siquiera logran dormirlo. Lo 'atontan' un poco y lo mantienen en un estado de reposo, pero que le permite, de vez en cuando, salir a 'pasear' por las células del organismo humano. Lo ha descubierto un equipo de investigadores españoles, que pone así fin a una de las controversias médicas más extendidas en el campo del sida.
Durante años, la teoría más apoyada ha sido la que consideraba que el cóctel de fármacos impedía la replicación del virus. No era capaz de eliminarlo, pero sí podía evitar que siguiera infectando. Otra hipótesis, más minoritaria, indicaba que a pesar de la terapia, el virus podía mantener algo de actividad. Y es esta última la que finalmente se ha confirmado, según la investigación que publica 'Nature Medicine' y que ha sido realizada por expertos del Instituto de Investigación del Sida IrsiCaixa, ubicado en el Hospital German Trias i Pujol de Badalona. Lo que han visto los investigadores, dirigidos por Javier Martínez Picado, es que algunos pacientes infectados por el VIH, a pesar de que presentan una carga viral indetectable gracias al tratamiento y que parecen tener la infección controlada, cuentan con una pequeña proporción del virus que sigue siendo infeccioso. Un descubrimiento que, según sus autores, "tiene importantes implicaciones clínicas, ya que podría explicar por qué, a pesar de que los pacientes permanecen en terapia durante muchos años, los fármacos no permiten curar la infección de manera definitiva". Martínez Picado lo aclara de forma sencilla a ELMUNDO.es: "Un paciente infectado sin tratamiento es como un bosque en llamas. Con los fármacos logramos apagar el fuego, pero quedan rescoldos y basta con un pequeño golpe de aire para que vuelva a incendiar. Esto es lo que hace el virus. Está apagado, pero no muerto". Carga viral indetectable Para llegar a estas conclusiones, el equipo analizó a 69 pacientes que seguían la terapia antirretroviral y que tenían carga viral indetectable (cantidad de virus en sangre) durante una media de cinco años. A 45 de ellos se les intensificó el tratamiento añadiendo otro fármaco, el Raltegravir, el primero de una nueva familia de medicamentos antisida, que bloquea la acción de la integrasa (una enzima que utiliza el VIH para copiar su material genético en el ADN de las células del paciente). Los otros participantes siguieron con su terapia antirretroviral normal.
La virología (rama de la microbiologia) es el estudio de los virus: su estructura, clasificación y evolución, su manera de infectar y aprovecharse de las células huésped para la reproducción del virus, su interacción con los organismos huéspedes, su inmunidad, la enfermedad que causan, las técnicas para su aislamiento, cultivo y su uso en investigación y terapia. La virología es considerada un sub-campo de la microbiología y la medicina. La virologia también es una rama taxonómica de la biología para estudiar los virus y todos sus componentes.
ESTRUCTURA DE LOS VIRUS
En los virus se distinguen las partes siguientes: 1.- Genoma vírico. Se compone de una o varias moléculas de ADN o de ARN, pero nunca los dos simultaneamente, Se trata de una sola cadena, abierta o circular, monocatenaria o bicatenaria. .
2.- Cápsida. Es la cubierta proteica que envuelve al genoma vírico. Está formada por proteínas globulares o capsómeros que se disponen de una manera regular y simétrica, lo que determina la existencia de varios tipos de cápsidas: icosaédricas, helicoidales y complejas.
La función de la cápsida es proteger el genoma vírico y, en los virus carentes de membrana, el reconocimiento de los receptores de membrana de las células a las que el virus parasita.
3.-Envoltura membranosa. Formada por una doble capa de lípidos que procede de las células parasitadas y por glucoproteínas incluidas en ella cuya síntesis está controlada por el genoma vírico. Las glucoproteínas sobresalen ligeramente de la envoltura y tienen como función el reconocimiento de la célula huesped y la inducción de la penetración del virus en ella mediante fagocitosis.
DIAPOSITIVA # 3 LA CÉLULA BACTERIANA Las bacterias son células muy sencillas; carecen de núcleo y tampoco presentan orgánulos en el citoplasma. Se las denomina Procariotas. Son organismos unicelulares y se encuentran en todos los ecosistemas. Las bacterias son un numeroso grupo de seres vivos, con características muy diversas. En la clasificación de los Dominios, Woese, aparecen dos grupos de Procariotas: el Dominio Archaea, que engloba a los organismos más antiguos del Planeta, y el Dominio Bacteria, en el que se encuentran la gran mayoría de los organismos bacterianos actuales, también conocidos con el nombre de Eubacterias. Las bacterias se caracterizan por no tener núcleo patente, ni clorofila típica, aunque pueden poseer otros pigmentos similares como bacterioclorina, bacterioclorofila y bacteriopurpurina, y sin pseudópodos. La bacterioclorina es un pigmento verde fotosintetizante que, según ciertos autores, acompaña a la bacteriopurpurina y a la bacterioclorifila en las algas purpúreas. La bacterioclorofila es otro pigmento fotosintetizante que, junto con la bacteriopururina, poseen las bacterias purpúreas. La bacteriopurpurina es un pigmento rojo que enmarca a la bacterioclorifila en las bacterias purpúreas, y que colabora con ella en la síntesis de los glúcidos, a partir del bióxido de carbono atmosférico. Esta síntesis se diferencia de la clorofílica en que utiliza el ácido sulfuhídrico como donador de hidrógeno. Aunque la organización de las bacterias es de tipo celular, el microscopio electrónico ha puesto al descubierto numerosas características propias de la célula bacteriana. También pueden ser observadas con el microscopio óptico. Una célula bacteriana se compone de una pared celular, membrana, citoplasma y ácido nucleico. La pared bacteriana aísla y protege perfectamente a la bacteria. Incluso algunas bacterias tienen una cápsula externa que las protege de los antibióticos y de los anticuerpos. La membrana bacteriana es esencialmente idéntica a la de las células eucariónticas, aunque posee unos entrantes en el citoplasma. En el citoplasma bacteriano las únicas estructuras existentes son los ribosomas y algunas vesículas llenas de gas. El ácido nucleico está formado por una sola cadena de ADN, que se suele llamar cromosoma bacteriano y es de forma circular, que se diferencia del cromosoma eucariótico en que es más pequeño y no se asocia tan íntimamente con las proteínas. Ambos se parecen en que se componen de ADN. Éste se halla condensado en una región de la bacteria llamada nucleoide o falso núcleo. Sus dimensiones son muy reducidas: tan sólo de algunas micras o fracción de micra. Se pensaba, no hace aún muchos años, que fuesen el límite inferior en la escala de ser vivientes; pero los modernos microscopios permiten observar otros seres mucho más diminutos, tales las rickettsias y los virus. Unas bacterias son inmóviles, otras poseen minúsculos flagelos, cuyo número y distribución varía notablemente, que les permiten desplazarse. Su capacidad reproductora es enorme, pues algunas se dividen cada 20 minutos si las condiciones les son favorables, por lo que una sola bacterias puede producir ingentes cantidades de descendientes en muy pocas horas. Se creyó durante mucho tiempo que sólo se reproducción asexualmente, pero hoy se conocen abundantes ejemplos se conjugación entre ellas, semejante a la de la reproducción sexual, al poder intercambiarse trozos de ADN, con lo que las bacterias resultantes tienen un material genético algo distinto. Se reproducen por bipartición simple, es decir, se parten en dos dividiendo equitativamente todo su contenido, incluido el ADN.
La base del diagnóstico viral es la detección del virus o de sus componentes. El aislamiento del virus era la técnica standard de oro sobre la cual se medían todas las otras pruebas de diagnóstico viral, pero hoy en día con el desarrollo de las nuevas técnicas de Biología Molecular ya no es la más sensible. De igual forma el aislamiento de virus tiene una sensibilidad y una especificidad muy alta. Debido a que sólo se amplifica el virus, se aumenta la sensibilidad sin disminuir la especificidad. Sin embargo, existen algunas desventajas en el aislamiento del virus: El proceso suele ser lento, ya que demanda días a semanas para la identificación, y en consecuencia puede no estar disponible a tiempo para influir en la atención del paciente. Además, es un proceso laborioso y caro. Por otra parte requiere el uso de sistemas de cultivos adecuados, por ejemplo, se necesitan varias líneas celulares para la detección óptima de virus.
Los cultivos celulares son, pues, los biosubstratos más corrientemente empleados para la propagación de los virus.
Un cultivo celular es obtenido, de explantes de órganos o de embriones de animales.
Estas células obtenidas asépticamente se disocian tratándolas con una enzima (tripsina) que rompe el cemento intercelular. La suspención de celulas libres así obtenidas se coloca en la superficie plana de un recipiente de vidrio o plástico en donde las células se adhieren y multiplican formando una fina capa de células que se llama MONOCAPA CELULAR.
Esta monocapa de células crece en un medio de cultivo complejo que contiene albúmina, vitaminas, sales, glucosa, etc., en un sistema buffer. Se previene la contaminación bacteriana adicionándole al medio antibióticos adecuados.
Los cultivos celulares en monocapa son los más usados, aunque hay otros sistemas (cultivos en suspensión, explantos, cultivos de órganos, cultivos en microcarriers, etc.).
Los cultivos celulares se dividen en:
Cultivos Primarios: Se obtienen a partir de células, tejidos u órganos tomados directamente del organismo y pueden subcultivarse una o dos veces.
Líneas Celulares Diploides: Son aquellas que crecen en pasajes sucesivos hasta aproximadamente 50 subcultivos y que conservan por lo menos en un 75% el cariotipo correspondiente a la especie de que provienen.
Líneas Celulares Continuas: Permite un número finito de subcultivos y son heteroploides. Para considerar que se ha logrado establecer una línea continua, esta debe de haber sido subcultivada por lo menos 70 veces. Las líneas celulares continuas ofrecen las siguientes ventajas:
Disponibilidad para todos los investigadores de stocks de células idénticas, ya sea congeladas en ampollas o en monocapa de botella de cultivos, con la posibilidad de reconstituirlas cuando sea necesario.
Facilidad relativa del pasaje y mantenimiento en todos los laboratorios.
Ciclo reproductivo de los virus. 1-Fijación 2-Penetración 3-Desenvolvimiento 4-Síntesis (4a-Transcripción, 4b-Traducción, 4c-Replicación del genoma) 5-Ensamblaje 6-Liberación El ciclo de replicación de los virus es el término utilizado para describir el ciclo de reproducción de los virus. Este consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden reproducirse por si mismos en el exterior; para ello requieren introducirse en células donde controlarán sus mecanismos reproductivos.
Virología es el estudio de virus y virus como agentes: clasificación y evolución, su estructura, sus formas de infectar y explotar las células para la reproducción del virus, las enfermedades que causan, las técnicas para aislar y la cultura de ellos y su uso en la investigación y terapia.
Virología es considerada una parte de la microbiología o de patología.
Una importante rama de la Virología es la clasificación de virus. Virus pueden clasificarse según la célula huésped que infectan:
virus animales, virus de plantas, hongos virus y bacteriófagos (virus que infectan bacterias, que son los virus más complejos).
Otra clasificación utiliza la forma geométrica de su cápside (a menudo una hélice o un icosaedro) o la estructura del virus (por ejemplo, la presencia o la ausencia de un envolvente de lípidos). Gama de virus en tamaño de unos 30 nm a aproximadamente 450 nm, lo que significa que la mayoría de ellos no puede verse con microscopios de luces.
El sistema de clasificación más útil y más ampliamente utilizado distingue virus segun el tipo de ácido nucleico que utilizan como material genético y el método de replicación viral que emplean coaxial células huésped en la producción de más virus:
• Virus de ADN (divididas en virus de ADN bicatenario y los virus de ADN monocatenario mucho menos común),
• Virus ARN (dividieron en virus de ARN monocatenario de sentido positivo, virus de ARN monocatenario de sentido negativo y los virus de ARN bicatenario mucho menos común),
• invertir transcribir virus (virus de ADN de transcripción inversa de bicatenario y monocatenario transcripción inversa RNA virus incluyendo retrovirus).
Un virus está compuesto de una molécula de ácido nucleico y una envoltura proteínica. Ésta es la estructura básica de un virus, aunque algunos de ellos pueden añadir a esto la presencia de alguna enzima, bien junto al ácido nucleico, como la transcriptasa inversa de los retrovirus, bien en la envoltura, para facilitar la apertura de una brecha en la membrana de la célula hospedadora.
A la unidad formada por el ácido nucleico y la envoltura proteínica se le denomina tambiénvirión.
El ácido nucleico es solamente de un tipo, ADN o ARN, nunca los dos.
Atendiendo al tipo de ácido nucleico se distinguen cuatro clases de virus:
- ADN de cadena doble
- ADN de cadena sencilla
- ARN de cadena doble
- ARN de cadena sencilla
La envoltura proteínica recibe el nombre de cápsida. Está formada por unas subunidades idénticas denominadas capsómeros. Los capsómeros son proteínas globulares que en ocasiones tienen una parte glicídica unida. Se ensamblan entre sí dando a la cubierta una forma geométrica.
Atendiendo la forma de la cápsida, se pueden distinguir los siguientes tipos de virus:
- Cilíndricos o helicoidales: los capsómeros, que son de un solo tipo, se ajustan entorno una hélice simple de ácido nucleico. Un ejemplo lo constituye el virus del mosaico del tabaco.
- Icosaédricos: los capsómeros, que suelen ser de varios tipos, se ajustan formando un icosaedro regular (es decir, 20 caras triangulares y 12 vértices), y dejando un hueco central donde se sitúa el ácido nucleico fuertemente apelotonado. Algunos forman poliedros con más caras que el icosaedro, y algunos presentan fibras proteicas que sobresalen de la cápsida.
Un ejemplo lo constituyen los adenovirus, entre los que se encuentran los virus de los resfriados y faringitis.
- Complejos: con pequeñas variantes, responden a la siguiente estructura general.
- Una cabeza de estructura icosaédrica que alberga el ácido nucleico. - Una cola de estructura helicoidal que constituye un cilindro hueco.
- Un collar de capsómeros entre la cabeza y la cola.
- Una placa basal, al final de la cola, con unos puntos de anclaje que sirven para fijar el virus a la membrana celular.
De la placa salen también unas fibras proteicas que ayudan a la fijación del virus sobre la célula hospedadora. Como ejemplo de este tipo de virus se encuentran la mayor parte de los virus bacteriófagos (que infectan las bacterias).
La simetría es la forma que adopta un virus en el espacio y está dad por la estructura de su nucleocápside. La simetría puede ser helicoidal, icosaédrica o compleja.
Los virus con simetría helicoidal presentan una nucleocápside cilindrica que puede estar extendida como en el virus del mosaico del tabaco. Este es un virus desnudo.
En otros casos la nucleocápside está arrollada sobre sí misma y recubierta por una envoltura, como en el virusInfluenza.
Los virus con simetría icosaédrica son poliedros regulares con 20 caras triangulares, 30 aristas y 12 vértices. Son ejemplos de virus con simetría icosaédrica desnudos el virus de la poliomielitis o losadenovirus. Los herpesvirus y togavirus son virus con simetría icosaédrica y recubiertos por una envoltura.
Se denominan virus de simetría compleja a aquellos que presentan una nucleocápside helicoidal o icosaédrica recubierta por una envoltura laxa. Ejemplos de éstos virus son los poxvirus (forma de ladrillo), los rhabdovirus (forma de bala) y algunos bacteriófagos que poseen una cabeza con simetría icosaédrica y una cola con simetría helicoidal.
Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus.
Su pequeño tamaño explica lo tardío del descubrimiento de estos agentes. La primera referencia sobre la existencia de los virus se debe al botánico ruso Dimitri Ivanovski en 1892. Este investigador buscaba el agente causante de la enfermedad denominada mosaico del tabaco, y llegó a la conclusión de que debía tratarse de una toxina o de un organismo más pequeño que las bacterias, pues el agente atravesaba los filtros que retenían las bacterias. Denominó a estos agentes patógenos virus filtrables.
En 1897, el microbiólogo holandés Martinus Beijerink realizó experimentos similares a los de Ivanovski, y llegó a desechar la idea de las toxinas, pues se trataba de un agente capaz de reproducirse, ya que mantenía su poder infeccioso de unas plantas a otras, sin diluirse su poder patógeno. Poco después, los microbiólogos alemanes Frederick Loeffler y Paul Frosch descubrieron que la fiebre aftosa del ganado era producida por un virus filtrable que actuaba como un agente infeccioso.
En la década de los 30, con el uso de filtros de tamaño de poro inferior, con las técnicas de cultivo celular in vitro que permitían la obtención de gran cantidad de virus, con la ultracentrifugación y finalmente con el microscopio electrónico y la difracción de rayos X, se logró visualizar a estos agentes.
Observaciones:
En la tabla correspondiente a esta diapositiva, disponemos un cuadro correspondiente al tamaño de cada uno de los cuales corresponden a esta categoría.
Nos muestras las diferentes familiar relacionadas a los virus las cuales conoceremos a continuación entre ellas están:
1- Picornaviridae: es una familia de virus infectivos para animales. Contienen un genoma ARN monocatenario positivo y por lo tanto se incluyen en el Grupo IV de la Clasificación de Baltimore. El genoma ARN es inusual porque tiene una proteína en el terminal 5' que se utiliza como iniciador de la transcripción por la ARN polimerasa. El nombre se deriva de "pico", que significa pequeño, con lo que "picornavirus" significa literalmente "virus RNA pequeños".
2- Caliciviridae: es una familia de virus infectivos para animales y causantes de gastroenteritis en humanos. Los calicivirus han sido encontrados en la mayoría de los animales domésticos y muchos silvestres, como cerdos, conejos, gallinas y anfibios. Contienen un genoma ARN monocatenario positivo y por lo tanto se incluyen en el Grupo IV de la Clasificación de Baltimore.
3- Hepadnaviridae es una familia de virus que causan infecciones en hígado de humanos y de animales. Los hepadnavirus tienen un genoma muy corto de ADN parcialmente de doble hélice y parcialmente de hélice simple, circular. Como su replicación involucra un ARN intermedio, se incluyen en el Grupo VII de la Clasificación de Baltimore.
4- Reoviridae es una familia de virus que pueden afectar el sistema gastrointestinal (tales como rotavirus ) y tracto respiratorio . Los virus de la familia Reoviridae han genomas que consiste en segmentar, ARN de doble cadena (dsRNA). El nombre "Reoviridae" se deriva de las vías respiratorias entéricas virus huérfanos.
5- Rhabdoviridae es una familia de virus infectivos para animales y plantas. Entre los animales infectados están insectos, peces y mamíferos, incluidos los humanos. Estos virus presentan una forma alargada parecida a una bala. Poseen un genoma de ARN monocatenario de sentido negativo por lo que se incluye en el Grupo V de la Clasificación de Baltimore.
6- El Togaviridae es una familia de virus que incluye los siguientes géneros: Género Alphavirus; especies típicas: Sindbis virus, Virus de la encefalitis equina oriental, Virus de la encefalitis equina occidental, Virus de la encefalitis equina venezolana, Virus del Río Ross, Virus O'nyong'nyong. Género Rubivirus; especies típicas: Rubéola virus.
7- Los Coronavirus son un género de virus ARN de vertebrados de Coronaviridae.1 Son virus envueltos con un genoma de ARN de cadena sencilla con polaridad positiva y simetría helicoidal. El tamaño genómico de los coronavirus varía entre 16 a 31 kilobases, un virus de extraordinaria longitud.
8- Bunyavirus, género de virus ARN de vertebrados de la familia Bunyaviridae.
9- Papovaviridae es una familia de virus pequeños (apróx. 50 nm) infectivos para una gran variedad de animales, incluyendo los humanos. Su nombre implica las patología que causa: papilomavirus + poliomavirus + agente vacuolante. Tienen una amplia distribución mundial.
10- Los paramixovirus son miembros de la familia de virus paramyxoviridae, que poseen un genoma con ARN de cadena sencilla de polaridad negativa como ácido nucleico; albergan dicha información genética en una cápside cubierta de envoltura viral y estructuralmente definida por una simetría helicoidal, de un tamaño de 125 a 250 x 18 nm, con envoltura, pleomórfico, y un filamentoso nucleocápside; y por ensamblar los viriones maduros en el citoplasma como compartimento celular.
11- Los Orthomyxoviridae son una familia de virus RNA que infectan a los vertebrados. Incluyen a los virus causantes de la gripe.
12- Un retrovirus es un virus de ARN que se replica en una célula huésped a través del proceso de transcripción inversa . En primer lugar, utiliza su propio transcriptasa inversa enzima para producir ADN de su ARN del genoma, la inversa de la pauta habitual, por lo tanto retro (hacia atrás).
13- Flavivirus : es un género de virus ARN pertenecientes a la familia Flaviviridae. Los Flavivirus son virus con envoltura, la simetría de la nucleocápside icosaedrica, y cuyo material genético reside en una única cadena de ARN de polaridad positiva.
14- Parvovirus, a veces truncada a "parvo", es a la vez el nombre común en Inglés casualmente se aplica a todos los virus de la Parvoviridae familia taxonómica y también el nombre taxonómico del parvovirus género dentro de la Parvoviridae familia.
15- Los adenovirus (Adenoviridae) son una familia de virus que infectan tanto humanos como animales. Son virus no encapsulados de ADN bicatenario que pueden provocar infecciones en las vías respiratorias, conjuntivitis, cistitis hemorrágica y gastroenteritis. Los adenovirus también se utilizan para obtener ADN para la terapia génica.
16- El Arenavirus es un género de virus de la familia Arenaviridae.La especie tipo es el virus de la coriomeningitis linfocítica (LCMV); es también de las spp. responsables de la fiebre Lassa. Arena, viene del latín con la familia de términos del árido.
17- Los herpesvirus o virus de la familia Herpesviridae, deben su nombre al término griego herpein (ἕρπειν), reptar o arrastrar, haciendo alusión a la facultad de estos agentes infecciosos microscópicos de ser fácilmente contagiados y transmitidos de una persona a la otra y de recurrencia crónica. Los herpesvirus forman una familia de virus divididos en tres subfamilias: alpha, beta y gammaherpesvirinae, ubicados con base en la arquitectura del virión y ciertas propiedades biológicas comunes.
18- Poxviridae es una familia de virus de ADN relacionados entre sí llamados poxvirus, infectivos para animales vertebrados (8 de los 11 géneros, por ejemplo, poxvirus de la gallina) y a los invertebrados (3 géneros). Tienen un genoma ADN bicatenario y por lo tanto pertenece al Grupo I de la Clasificación de Baltimore.
19- Filoviridae es una familia de virus del orden Mononegavirales. Se caracterizan por poseer como genoma RNA monocatenario de sentido negativo, y por infectar a primates. Agrupa, entre otros, al virus Ébola y al virus Marburg.
Los virus son organismos tan pequeños, que su tamaño les permite pasar a través de los filtros que retienen bacterias. Las características básicas para identificar a un virus son: la composición del ácido nucleico, la disposición de la cápside, la presencia o ausencia de una envoltura, el tamaño del virus y el huésped.
MUESTRAS PARA DIAGNOSTICO:
Las muestras deben tomarse precozmente en la fase aguda de la infección. Deben transladarse al laboratorio a la mayor brevedad posible, en medio de transporte. Las muestras más comunes son:
• Torundas nasofaríngeas: adecuados para el diagnóstico de enterovirus, adenovirus, y herpes simplex. Las nasofaringeas se prefieren para el virus respiratorio sincicial y la mayoria de los otros respiratorios. Las nasales son las mejores para los rinovirus.
• Aspirados faríngeos: son mejores que los hisopos (torundas) nasofaringeos, pero estos últimos son mas prácticos.
• Torundas rectales y muestras de heces: Aquí se muestran virus no cultivables como rotavirus, o adenovirus, que se detectan por microscopía electrónica o con técnicas de detección de antígenos.
• Secreciones, Sangre o Líquido cefalorraquídeo.
• En algunas virosis excepcionalmente el diagnóstico solo puede efectuarse en Biopsia de cerebro, como en la encefalitis por herpes simplex; en Huéspedes inmunodeprimidos y con citomegalovirus, el virus solo puede identificarse en el órgano afectado.
DIAGNOSTICO VIRAL
Observación del efecto citopático, en cultivo de tejidos inoculados con ciertos virus. La etiología de un determinado síndrome viral puede demostrarse recuperando el agente causal en tejidos celulares de diversos tipos (por ej: riñón de mono, pulmón humano fetal, amnios humano o células tumorales humanas). Los huevos embrionados actualmente solo se utilizan para cultivar el virus cuando se desea preparar vacunas. El empleo de animales de experimentación es excepcional en el diagnóstico.
• Las células primarias de riñón de mono son excelentes para cultivar mixovirus, paramixovirus y muchos enterovirus; también para algunos adenovirus.
• Las células diploides fetales humanas son células fibroblásticas en las que crece un amplio espectro de virus, como el de herpes simple, varicela zoster, adenovirus, picornavirus, y sólo en éstas pueden cultivarse los citomegalovirus.
• Las células HEP-2 son epiteliales provenientes de un cancer humano, útiles para recuperar el virus respiratorio sincitial, adenovirus, y herpes simple.
• Células mononucleares normales normales humanas de sangre periférica, que no suelen estar disponibles, permiten el cultivo del virus ( SIDA ) de la inmunodeficiencia, el coxsackie A y los togavirus.
El crecimiento de un virus se detecta mediante la observación de cambios en la monocapa del cultivo celular, denominado efecto citopático (ECP). Cuando aparecen cambios en la morfología celular, lisis celular, vacuolización, formación de cincicios y existen cuerpos de inclusión, se produce un efecto citopático característico. Los cuerpos de inclusión son cambios histológicos que se producen en las células por los componentes virales o por variaciones en las estructuras celulares.
Un técnico experto puede distinguir los efectos citopáticos característicos de las principales familias de virus. Según el cultivo celular en el que aparece el ECP y el tipo de crecimiento viral, se puede realizar un diagnóstico de presunción de un gran número de virus de importancia clínica.
Algunos virus no producen rápidamente ECP en las líneas celulares que se suelen utilizar en los laboratorios de virología clínica. No obstante se pueden determinar por otras técnicas, como la interferencia en la replicación de otros virus (por ej los picornavirus no pueden replicarse en células previamente infectados con el virus de la rubeola, lo que se denomina interferencia heteróloga).
1.2. Fijación de eritrocitos en cultivos celulares infectados con virus. En el caso de virus que dan lugar a este fenómeno, como mixovirus o virus de la parotiditis, se observa una hemaglutinación al adicionar eritrocitos a la monocapa de células.
1.3. Detección de antígenos virales. Cuando los virus se pueden detectar antes de desarrollar el efecto citopático, se utilizan anticuerpos monoclonales dirigidos contra antígenos tempranos y técnicas de inmunofluorescencia o inmunoperoxidasa.
1.4. Inmunofluorescencia directa, utilizando células del paciente, como leucocitos, orina o mucosa oral o genital, para buscar virus.
1.5. Detección de ácidos nucleicos virales por reacción de polimerasa en cadena, las cuales permiten amplificar una porción pequeña de ácido nucleico en un período de tiempo corto.
1.6. Microscopía electrónica. Cuando algunos virus comunes no se aislan con los métodos de cultivo habituales, ni pueden diagnosticarse por técnicas serológicas, pueden estudiarse con otras técnicas, como la microscopía electrónica. Tal es el caso de los calcivirus, los minirrotavirus, los astrovirus y los parvovirus del cerebro. Sin embargo, hay tres grandes dificultades para su uso: el instrumento es caro y es difícil su mantenimiento. Por otro lado, deben estar presentes grandes cantidades de particulas virales en la muestra para poder detectarse. Finalmente, la morfología no es suficientemente diferencial para un diagnóstico preciso.
4-Síntesis (4a-Transcripción, 4b-Traducción, 4c-Replicación del genoma)
5-Ensamblaje
6-Liberación
El ciclo de replicación de los virus es el término utilizado para describir el ciclo de reproducción de los virus. Este consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden reproducirse por si mismos en el exterior; para ello requieren introducirse en células donde controlarán sus mecanismos reproductivos.
La adhesión es la propiedad de la materia por la cual se unen y plasman dos superficies de sustancias iguales o diferentes cuando entran en contacto, y se mantienen juntas por fuerzas intermoleculares.
Mecanismos de adhesion
La cohesión es la causa de que el agua forme gotas, la tensión superficial hace que se mantengan esférica y la adhesión la mantiene en su sitio.
Las gotas de agua son más planas sobre la flor de Hibiscus ya que tienen mejor adhesión.
Cinco mecanismos han sido propuestos para explicar por qué un material se adhiere a otro.
Adhesión mecánica
Los materiales adhesivos rellenan los huecos o poros de las superficies manteniendo las superficies unidas por enclavamiento. Existen formas a gran escala de costura, otras veces a media escala como el velcro y algunos adhesivos textiles que funcionan a escalas pequeñas. Es un método similar a la tensión superficial.
Adhesión química
Dos materiales pueden formar un compuesto químico al unirse. Las uniones más fuertes se producen entre átomos donde hay permutación (enlace iónico) o se comparten electrones (enlace covalente). Un enlace más débil se produce cuando un átomo de hidrógeno que ya forma parte de una partícula se ve atraída por otra de nitrógeno, oxígeno o flúor, en ese caso hablaríamos de un puente de hidrógeno.
Adhesión dispersiva
En la adhesión dispersiva, dos materiales se mantienen unidos por las fuerzas de van der Waals: la atracción entre dos moléculas, cada una de las cuales tiene regiones de carga positiva y negativa. En este caso, cada molécula tiene una región de mayor carga positiva o negativa que se une a la siguiente de carga contraria. Este efecto puede ser una propiedad permanente o temporal debido al movimiento continuo de los electrones en una región.
En la ciencia de superficies el término "adhesión" siempre se refiere a una adhesión dispersiva. En un sistema sólido-líquido-gas normal (como una gota de un líquido sobre una superficie rodeada de aire) el ángulo de contacto es usado para cuantificar la adhesividad. En los casos donde el ángulo de contacto es bajo la adhesión está muy presente. Esto se debe a que una mayor superficie entre el líquido y el sólido conlleva una energía superficial mayor.
Adhesión electrostática
Algunos materiales conductores dejan pasar electrones formando una diferencia de potencial al unirse. Esto da como resultado una estructura similar a un condensador y crea una fuerza electrostática atractiva entre materiales. mi con diversos materiales les pueden ayudar.
Los virus son moléculas de DNA o RNA rodeadas por una envoltura proteica que necesitan células viables para poder replicarse. Los virus utilizan la maquinaria metabólica de las células para sintetizar su material genético y proteínas de la envoltura. Existen distintos tipos de virus que pueden infectar células procariontes o células eucariontes. Los bacteriófagos o fagos son virus que se reproducen en células procariontes.
El genoma de los fagos puede ser RNA simple cadena (MS2, Qß), RNA doble cadena (phi 6), DNA simple cadena (phi X174, fd, M13) o DNA doble cadena (T3, T7, lambda , T5, Mu, T2, T4). Estos ácidos nucleicos pueden contener bases inusuales que son sintetizadas por proteínas del fago. En los T-pares el genoma no contiene citosina sino 5'-
hidroximetilcitosina, mientras que en otros tipos de fago alguna de las bases esta parcialmente sustituida.
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante.
Maduración y ensamblaje
Cuando se han sintetizado un número crítico de varios componentes virales, éstos se ensamblan en partículas víricas maduras en el núcleo y/o citoplasma de la célula infectada. En este ensamblaje y al contrario de lo que ocurre en los bacteriófagos, no están implicados enzimas biosintéticos especiales, sino que ocurre de forma espontánea debido a la gran afinidad de la cápsida con el ácido nucleico viral. La maduración se puede definir como la fase de la infección viral durante la cual los componentes estructurales del virión se producen y ensamblan con el nuevo ácido nucleico viral para formar la nucleocápsida.
#12 Maduración y Salida de Virus Desnudos y Envueltos
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. (Figura 1).
(B) Entada vía endosomas en la superficie celular (figura 2) Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Los antivirales son un tipo de medicamento usado para el tratamiento de infecciones producidas por virus. Tal como los antibióticos (específicos para bacteria), existen antivirales específicos para distintos tipos de virus. No sin excepciones, son relativamente inocuos para el huésped, por lo que su aplicación es relativamente segura. Deben distinguirse de los viricidas, que son compuestos químicos que destruyen las partículas virales presentes en el medio ambiente.
El primer antiviral exitoso, el aciclovir, es un análogo a los nucleósidos y es efectivo contra el herpesvirus. La primera sustancia antiviral en ser aprobada para tratar el VIH, la zidovudina (AZT), es también un nucleósido análogo.
Un mejor conocimiento de la acción de la transcriptasa inversa ha conducido a mejores análogos a nucleósidos para tratar las infecciones del VIH. Uno de estos medicamentos, la lamivudina, ha sido aprobado para tratar la hepatitis B, que utiliza la transcriptasa inversa como parte de su proceso de replicación. Los investigadores han ido más lejos y han desarrollado inhibidores que no parecen nucleósidos, pero pueden seguir bloqueando la transcriptasa inversa.
Otros objetivos a ser considerados por los antivirales VIH incluyen la RNasa H, que es un componente de la transcriptasa inversa que rompe el ADN sintetizado del ARN viral original y la integrasa, que empalma el ADN sintetizado con el genoma de la célula huésped.
Los retrovirus son pequeños microorganismos que contienen una única cadena de ARN. Su ciclo vital se basa en la conversión de su ARN geonómico en doble cadena de ADN y posteriormente en la integración de éste último en los cromosomas de la célula huésped en lo que se denomina provirus endógeno. De forma ocasional algunas secuencias celulares son recombinadas con el provirus y a su vez estas recombinaciones pueden transferirse a otra población celular infectada, lo que puede tener importantes consecuencias en las funciones celulares.
Peyton Rous identificó el primer retrovirus en 1911 cuando se encontró un agente transmisible capaz de causar un tipo de sarcoma en pollos. Fue tras el descubrimiento del enzima transcriptasa inversa por Temin cuando se pudo ahondar en sus mecanismos de acción y en su manipulación experimental.
Como se comentó al inicio de esta exposición fue hacia 1976 cuando se demostró que los supuestos oncogenes que portaban algunos de estos virus tenían una contrapartida en humanos como genes normales a los que se denominaron protooncogenes.
Otro elemento esencial en el estudio de los retrovirus lo constituyó el descubrimiento del primer retrovirus infeccioso humano el HTLV-1 al que luego se le sumaría el HIV.
La familia de los retrovirus ha sido subclasificada en tres subfamilias: Oncornavirus (que incluye al virus del tumor mamario murino, el virus de la reticuloendoteliosis aviar, virus de la leucemia humana de células T), Lentivirus (que incluye los virus de las inmunodeficiencias) y los Spumavirus.
Retroviridae es una familia de virus que comprende los retrovirus. Son virus con envoltura que presentan un genoma de ARN monocatenario de polaridad + y se replican de manera inusual a través de una forma intermedia de ADN bicatenario.
Este proceso se lleva a cabo mediante una enzima: la retrotranscriptasa o transcriptasa inversa, que dirige la síntesis de ADN a través de ARN y posee una importancia extraordinaria en la manipulación genética. Una vez se ha pasado de ARN monocatenario a ADN se inserta dentro del ADN propio de la célula infectada donde se comporta como un gen más (véase Ciclo reproductivo de los virus). Por tanto, se incluyen en el Grupo VI de la Clasificación de Baltimore.
Los retrovirus son responsables de muchas enfermedades, incluyendo algunos cánceres y el sida (VIH). Existen diversos grupos de investigación que han intentado modificar genéticamente los retrovirus para usarlos en terapia génica como vectores, pero se han encontrado con diversos problemas.
Virus: es un agente infeccioso microscópico que sólo puede multiplicarse dentro de las células de otros organismos.Los virus infectan todos los tipos de organismos, desde animales y plantas, hasta bacterias y arqueas.
Los virus son demasiado pequeños para poder ser observados con la ayuda de un microscopio óptico, por lo que se dice que son submicroscópicos; auque existen excepciones entre los Virus nucleocitoplasmáticos de ADN de gran tamaño, tales como el Megavirus chilensis, el cual se logra ver a traves de microscopía óptica.
Un virusoide o ARN satélite es una pequeña molécula patógena de ARN (ácido ribonucleico) que se encuentra encapsulada en ciertos virus de plantas con secuencias de ARN de mayor tamaño, lo que se denomina un virus asistente, el cual hospeda al virusoide y es necesario para que éste produzca una infección exitosa.
Los viroides son agentes infecciosos que, al igual que los virus, tienen un ciclo extracelular que se caracteriza por la inactividad metabólica y un ciclo intracelular en el que causan infección al huésped susceptible, pero que a diferencia de los virus, los viroides no poseen proteínas ni lípidos y están constituidos por una cadena cíclica corta de ARN, circular o con forma de varilla, (que no codifica proteínas). Es importante decir que tanto su forma intracelular como extracelular son las mismas (ARN desnudo), los mecanismos por los cuales éstos logran causar infección están relacionados con la autocatálisis de su material genético.
Los priones son los agentes causantes de un grupo de patologías neurodegenerativas letales características de mamíferos, también conocidas como encefalopatías espongiformes transmisibles. Estos agentes son capaces de propagarse dentro de un mismo huesped causando una lesión espongiótica y de transmitirse de huesped a huesped con elevados tiempos de incubación.
Transposon: Secuencia de DNA que puede moverse de un lugar a otro del cromosoma, insertar copias adicionales de ella misma en otros puntos o pasar de un cromosoma a otro. Tramo de DNA que puede incorporarse en otras moléculas de DNA en lugares donde no hay homología de secuencia. Tienen un tamaño entre 1 y 40 kb. Codifican todas las enzimas necesarias para su inserción. Pueden desplazarse dentro de un cromosoma o entre cromosomas.
Los plásmidos son moléculas de ADN extracromosómico circular o lineal que se replican y transcriben independientes del ADN cromosómico. Están presentes normalmente en bacterias, y en algunas ocasiones en organismos eucariotas como las levaduras. Su tamaño varía desde 1 a 250 kb.
Los viroides, al igual que los virus son parásitos intracelulares estrictos, es decir, dependen de la célula que infectan para su multiplicación, pero aquí acaban las similitudes. El virus más pequeño tiene un genoma de unas 1500 bases. En cambio el genoma de un viroide está comprendidos entre 250 y 400 nucleótidos (unas 20 veces más pequeños). El genoma de los virus puede ser de ADN o de ARN de doble hebra o de cadena simple, y a pesar que parasitan la maquinaria celular del huésped, siempre codifican alguna proteína. Los viroides en cambio siempre son estructuras de ARN de una sola hebra, circulares y su secuencia no codifica ninguna proteína. Los virus tienen una cápsida o cobertura, los viroides en cambio son la hebra de ARN desnuda.
Los viroides fueron descubiertos en los años 1970 por Theodore Otto Diener, al demostrar que el agente de la enfermedad del tubérculo fusiforme de la patata era una entidad más pequeña que los virus. La comunidad científica se mostró reacia al descubrimiento y en un principio se achacó a que pudiera ser un ARN satélite (secuencias de ARN que parasitan a otros virus… si, algunos virus también tienen sus propios parásitos). Pero no era el caso. Y quizás esto explica por que se habla tan poco de ellos en los libros de biología. Solo existen en plantas. No existen viroides en animales, aunque el virus de la hepatits delta tiene ciertas similitudes con los viroides, no se ha podido demostrar un origen evolutivo común.
Los viroides son los agentes infecciosos de menor complejidad genética y estructural conocidos y representan una forma extrema de parasitismo. Están constituidos exclusivamente por moléculas de ARN de cadena simple, cerradas covalentemente o con forma de bastón de bajo peso molecular (246 a 1994 nucleótidos). Carecen de actividad de ARN mensajero y se replican de forma autónoma, utilizando el sistema de transcripción de la célula suceptible.2 Los viroides se encuentran, casi exclusivamente, en el núcleo de las células infectadas; se desconoce el modo en que se replican pero se sabe que el ARN que los constituyen no funciona como ARN mensajero y tampoco se traduce a enzimas que participen en su propia replicación. Los viroides no codifican para ninguna proteína y llevan a cabo su replicación en la célula hospedadora a través de la RNA polimerasa II codificada por la célula hospedadora aunque también se ha propuesto la posibilidad de que lo haga gracias a una RNA polimerasa dependiente de RNA presente en algunas células eucariotas (Cann, 2001). Dada su localización, se presume que causan la enfermedad interfiriendo con la regulación génica de la célula hospedadora en la etapa de corrección del ARN mensajero celular: particularmente en la eliminación de intrones y de empalmes de exones, presentando secuencias muy similares a los intrones que normalmente son cortados. Sus ARN se replican varias veces.
Un virus satélite es un agente subvírico formado por ácido nucleico y que depende de la coinfección de la célula huésped por un virus (helper o máster) para reproducirse; obteniendo de ellos lasenzimas faltantes en su genoma, las cuales son necesarias para lograr replicarse.
Un virusoide o ARN satélite es una pequeña molécula patógena de ARN (ácido ribonucleico) que se encuentra encapsulada en ciertos virus de plantas con secuencias de ARN de mayor tamaño, lo que se denomina un virus asistente, el cual hospeda al virusoide y es necesario para que éste produzca una infección exitosa.
La McIntosh Roja o Mac es un tipo de manzana de piel roja y verde. Se recoge a finales de septiembre y es el cultivo más popular de Nueva Inglaterra, muy conocida por la compota rosada que se hace a partir de manzanas McIntosh sin pelar. Cada manzana de McIntosh tiene un linaje directo a un solo árbol descubierto en 1796 por John McIntosh en su granja en Dundela, una aldea situada en el condado de Dundas en la provincia canadiense de Ontario, cerca de Prescott.
Los plásmidos son fragmentos extracromosómicos de ácidos nucléicos (ADN o ARN) que aparecen en el citoplasma de algunos procariotas. Son de tamaño variable aunque menor que el cromosoma principal.
Los priones son los agentes causantes de un grupo de patologías neurodegenerativas letales características de mamíferos, también conocidas como encefalopatías espongiformes transmisibles. Estos agentes son capaces de propagarse dentro de un mismo huesped causando una lesión espongiótica y de transmitirse de huesped a huesped con elevados tiempos de incubación. A diferencia de virus y viroides, son resistentes a tratamientos inactivantes de ácidos nucléicos, pero comparten con éstos la existencia de una variabilidad de inóculos dentro de la misma especie (diferenciables por el patrón de la lesión y la magnitud del tiempo de incubación) y de una infectividad sujeta a barrera de especie (Chandler, 1961; Alper y cols., 1967; Hunter, 1972; Prusiner, 1982; Bruce y Fraser, 1991; Bessen y Marsh, 1992).
La búsqueda de la entidad molecular constitutiva de este agente reveló como componente mayoritario, si no único, una proteína: PrPSc, proteína del prion de scrapie),y la ausencia de un ácido nucleico específico (Prusiner 1982, 1991). Con estas premisas estructurales unidas a la capacidad de infección, Prusiner acuña el término prion (particula infecciosa de naturaleza proteica) para diferenciarlo de virus y viroides. Dadas las características poco convencionales de los priones se han elaborado numerosas hipótesis sobre su estructura (Dickinson y Outram, 1988; Prusiner, 1991; Weissmann, 1991). En la actualidad la hipótesis con mayor grado de aceptación es la conocida como "sólo proteína", delineada inicialmente por Griffith (1967) y, formalmente enunciada y actualizada por Prusiner (1991, 1997). Tras la descripción de proteínas de diferente secuencia pero similar comportamiento en levadura así como los avances interdisciplinares realizados en el conocimento de estos agentes en la última década, el concepto de prion ha sido acotado adquiriendo una defición más precisa y generalizable (Wicker y Masison, 1996; Lindquist S., 1997; Prusiner, 1997). Así, se denomina prion a la forma alterada de una proteína celular funcional (PrP en mamíferos) que ha podido perder su función normal pero que ha adquirido la capacidad de transformar la forma normal en patológica.
Han sido definidos como pequeñas partículas proteináceas que son resistentes a la inactivación por procedimientos que modifican los ácidos nucléicos.
Otra definición de prion es la forma alterada de una proteína celular funcional que ha podido perder su función normal pero que ha adquirido la capacidad de transformar la forma normal en patológica.
Los mamíferos tienen en su genoma genes que codifican la “proteína priónica”(PrP), y esta proteína en condiciones normales tiene una configuración espacial de tipo hélices alfa, que es susceptible a la lisis por proteasas y no es patógena. Los priones no son seres vivos ni tienen la capacidad de infección como tal.
La patogénesis de estas proteínas se produce cuando tienen bien una mutación (esporádica o adquirida genéticamente) o bien un cambio conformacional inducido por PrP-Sc (patógena) de otro individuo.
El cambio conformacional o las mutaciones hacen que la PrP tenga una estructura tridimensional diferente constituida por hélices beta con agregados amorfos insolubles (PrP-Sc), de manera que se hace imposible su lisis por las proteasas, produciéndose su acúmulo en los lisosomas de las células de los tejidos afectados y originando así las enfermedades priónicas.
Hugo Avila Lamarche(2011-0232)(Grupo No.3) La virología (rama de la microbiologia) es el estudio de los virus: su estructura, clasificación y evolución, su manera de infectar y aprovecharse de las células huésped para la reproducción del virus, su interacción con los organismos huéspedes, su inmunidad, la enfermedad que causan, las técnicas para su aislamiento, cultivo y su uso en investigación y terapia. La virología es considerada un sub-campo de la microbiología y la medicina. La virologia también es una rama taxonómica de la biología para estudiar los virus y todos sus componentes.
Su investigación incluye:
* la replicación viral * los patógenos virales. * la inmunología viral. * las vacunas virales. * los métodos de diagnóstico. * la quimioterapia antiviral. * las medidas de control de una infección. * los diferentes signos que manifiestan la presencia de virus.
yuleika veloz 2011-0481 Las células eucariotas surgieron de antepasados procariotas. El origen de los eucariotas se encuentra en sucesivos procesos simbiogenéticos (procesos simbióticos que culminan en la unión de sus simbiontes, estableciéndose una nueva individualidad de los integra) entre diferentes bacterias.
La teoría endosimbiótica postula que algunos orgánulos propios de las células eucariotas, especialmente plastos y mitocondrias, habrían tenido su origen en organismos procariotas que después de ser englobados por otro microorganismo habrían establecido una relación endosimbiótica con éste. Se especula con que las mitocondrias provendrían de protebacterias alfa (por ejemplo, rickettsias) y los plastos de cianobacterias.
Mas tarde con el tiempo y por evolución celular estas bacterias quedarían transformadas en Mitocondrias (bacterias aerobias) y en Cloroplastos (bacterias Fotótrofas), no tiene que ver con la evolución de las algas sino con la aparición de Mitocondrias y cloroplastos, los 2 organelos membranosos que se encuentran únicamente en células eucariotas poseen ADN de tipo procariota, ARN y Ribosomas dentro del mismo, pero la Teoría Endosimbiótica es mas aplicada en las Mitocondrias porque aparte de poseer 1sola molécula de ADN de tipo Procariota, ARN y Ribosomas en el lado interno de las "Crestas Mitocondriales que son proyecciones incompletas hacia la Matriz mitocondrial de la membrana interna" esta la Cadena Oxidativa o respiratoria.
Estas se diferencian principalmente, en que mayoría de las células eucariotas se distinguen de las procariotas por la existencia, en su citoplasma, de varios millares de estructuras organizadas, los orgánulos, de cuyo tamaño viene a ser el de una célula procariota.
Célula Bacteriana:
Las células bacterianas esta constituida principalmente, por elementos facultativos y obligados.
Elementos obligatorios : están presentes en todas las bacterias y son indispensables para la vida de la propia bacteria estos son los siguientes: - Pared celular - Membrana citoplasmática - Citoplasma - Ribosomas - Región nuclear Elementos facultativos: pueden estar o no presentes en la bacteria y son: - Cápsula - Flagelos - Fimbrias o Pili(pelo) - Endosporas - Inclusiones citoplasmáticas
La gram positiva mide de unos 15 a 20 nanometros de diametro y la gram negativa mide 2 nanometros de diametro.
La pared gram positiva, poseen una pared celular interna y una pared de peptidoglucano(es un exoesqueleto que da consistencia y forma esencial para replicación y supervivencia de la bacteria), no tiene membrana externa, no tiene espacio periplasmático, la red de mureína está muy desarrollada y llega a tener hasta 40 capas, conservan el complejo yodocolorante.
La pared gram negativa, poseen una pared celular más compleja: pared celular interna, pared de peptidoglucano, bicapa lipídica externa, tiene membrana externa que forma un saco rígido alrededor de la bacteria, mantiene estructura y es barrera impermeable a macromoléculas, ofrece protección en condiciones adversas, tiene espacio periplasmático, pueden ser anaerobios o aerobios y poseen proteínas con concentraciones elevadas.
Coloración del Gram
La tinción de Gram o coloración de Gram es un tipo de tinción diferencial empleado en Bacteriología para la visualización de bacterias, sobre todo en es una prueba potente y rápida que nos permite diferenciar dos clases de bacterias.
Es una prueba potente y rápida que nos permite diferenciar dos clases de bacterias estas son: Bacterias gram positivas y las gram negativas.
Las grampositivas se tiñen de morado ya que el colorante se queda atrapad en la capa gruesa de peptidoglucanos que rodea a la célula
Las gramnegativas tienen una capa de petidoglucano mucho más delgada es por ello que no retiene el violeta cristal y por esto las células se tiñen con safranina y las observamos roja
Formas y agrupaciones bacteriana
Los principales tipos de formas bacterianas son:
*Cocos (células más o menos esféricas)
*Bacilos (en forma de bastón, alargados),que a su vez pueden tener varios aspectos:cilíndricos,fusiformes y en forma de maza, etc.
Atendiendo a los tipos de extremos, éstos pueden ser:
*Redondeados (lo más frecuente)
*Cuadrados
*Biselados
*Afilados
*Espirilos: al igual que los bacilos, tienen un eje más largo que otro, pero dicho eje no es recto, sino que sigue una forma de espiral, con una o más de una vuelta de hélice.
*Vibrios: proyectada su imagen sobre el plano tienen forma de coma, pero en el espacio suelen corresponder a una forma espiral con menos de una vuelta de hélice.
Otros tipos de formas son filamentos, ramificados o no,anillos casi cerrados, y formas con prolongaciones (con prostecas).
2011-0481 FAMILIAS DE VIRUS. -PicoRNAvirus: Por picornavirus se conoce a todo virus que pertenece a la familia Picornaviridae. Los picornavirus son virus no envueltos, de única cadena ARN con cápside icosaédrica.
Su genoma es inusual ya que tiene una proteína en el extremo 5 ‘que se utiliza como cebador para la transcripción por la ARN polimerasa. El nombre se deriva de “pico”, que significa pequeño, ya que es literalmente un pequeño virus de ARN.
Los picornavirus se separan en una serie de géneros e incluyen muchos importantes patógenos de humanos. Las enfermedades que causan son muy variadas, que van desde aguda “gripe común” a enfermedades como la poliomielitis, y otras infecciones crónicas. - Los Calicivirus: Son Virus RNA pequeños pertenecientes a la familia Caliciviridae. Dentro de esta familia destacan dos géneros: los Norovirus, también conocidos como "virus tipo Norwalk" o "virus pequeños redondos estructurados" y los Sapporovirus. - Hepadnavirus: Los hepadnavirus son un subgrupo de los virus que producen hepatitis. Es uno de los virus más importantes desde el punto de vista médico. Infecta a más de un cuarto de billón de personas y es una de las principales causas de cirrosis y cáncer hepático (carcinoma hepatocelular). HBV, también representa el primer ejemplo del éxito de una vacuna recombinante para una enfermedad contagiosa humana, y puede tener importantes implicaciones para otras enfermedades de transmisión sexual, como el HIV. - Reovirus: Grupo de tres virus de doble cadena de ARN presentes en todas partes, identificados en los aparatos respiratorio y digestivo tanto de personas sanas como de enfermas. Los reovirus han sido implicados en algunos casos de enfermedades de vías respiratorias superiores y de gastroenteritis infantiles. - Los Rhabdovirus: Son virus con cápside helicoidal. Incluye al virus de la rabia y de la estomatitis vesicular (que es el organismo modelo). El virus de la estomatitis vesicular es un virus de 180x75nm, con cápsida rodeada por una envoltura con proyecciones constituídas por la proteína G. - Togavirus: Familia de arbovirus a la que pertenecen los organismos causantes de encefalitis, del dengue, de la fiebre amarilla y de la rubéola. - Bunyavirus: Género de virus ARN de vertebrados de la familia Bunyaviridae. -Papovavirus: Grupo de pequeños virus ADN, algunos de los cuales pueden ser potencialmente productores de cáncer. La verruga humana está producida por un tipo de papovavirus. Algunos tipos de papovavirus son: papovavirus del papiloma, papovavirus del polioma y papovavirus SV. - Paramixovirus: Miembro de una familia de virus que abarca a los organismos causantes de la gripe, de las paperas y de algunas infecciones respiratorias. - Retrovirus: Virus de ácido ribonucleico que posee una enzima que le permite convertir su material genético en ácido desoxirribonucleico cuando ha penetrado en una célula viva. Al grupo de los retrovirus pertenece el virus del sida. - Filovirus: Género de virus con ácido ribonucleico de cadena simple con sentido negativo, de la familia Filoviridae. El género incluye los virus Ébola y Marburg. - Poxvirus: Miembro de una familia de virus que incluye a los organismos que producen el molusco contagioso, la viruela y la vacuna.
*DIAGNOSTICO DE LAS VIROSIS. ¿Cómo infectan los virus al ser humano? Los virus pueden penetrar en el cuerpo humano por cualquiera de sus “puertas de entrada,” pero por lo general entran por la nariz y la boca. Una vez dentro del cuerpo, el virus se adhiere a la superficie de la célula que piensa atacar, denominada célula huésped. Una vez dentro, comienza a elaborar otros virus idénticos a partir de la proteína de la célula huésped. Los nuevos virus pueden salir a través de la membrana externa de la célula huésped, a la que a veces destruyen antes de abandonarla, y se lanzan al ataque contra otras células huésped. El proceso prosigue hasta que el cuerpo elabora suficiente cantidad de anticuerpos y otras defensas para derrotar a los virus invasores.
El término acelular significa literalmente "sin células". Se aplica principalmente a los virus, pero también a provirus y viroides, que son entidades biológicas no formadas por células, a diferencia de bacterias, compuestas por células procariotas, y de hongos, vegetales y animales, formados por células eucariotas.
LOS VIROIDES I y II
son agentes infecciosos que, al igual que los virus, tienen un ciclo extracelular que se caracteriza por la inactividad metabólica y unciclo intracelular en el que causan infección al huésped susceptible, pero que a diferencia de los virus, los viroides no poseen proteínas ni lípidos y están constituidos por una cadena cíclica corta de ARN, circular o con forma de varilla, (que no codifica proteínas).1 Es importante decir que tanto su forma intracelular como extracelular son las mismas (ARN desnudo), los mecanismos por los cuales éstos logran causar infección están relacionados con la autocatálisis de su material genético. Estas pequeñas moléculas de ARN circular de cadena sencilla formadas por unos pocos cientos de nucleótidos. Carecen de cápsida y su tamaño es una milésima parte del de los virus más pequeños. - Aparecen con mayor abundancia en el núcleo de las células infectadas. - Sólo han sido detectados en plantas, en las que producen una gran variedad de enfermedades y pueden ser transmitidos tanto horizontal como verticalmente. - No existen ninguna evidencia de que los viroides son traducidos a proteínas, ni se conoce cómo causan las enfermedades. Lo que sí se sabe es que son replicados por los enzimas del huésped. - Algunas semejanzas entre la secuencia de nucleótidos de los viroides y ciertos secuencias del genoma de las plantas no traducidas (intrones) sugieren que los viroides evolucionaron a partir de los intrones.
Actualmente se dispone de farmacos antivirales de probada eficacia terapeutica y profilactica para varias infecciones virales comunes y algunas potencialmente mortales. Ademas, continua la busqueda de nuevos farmacos antivirales y nuevas estrategias terapeuticas para el tratamiento de las enfermedades virales. Se pueden clasificar en tres grandes grupos a los agentes quimioterapicos activos frente a los virus:
Farmacos Viricidas: farmacos que directamente inactivan los virus intactos. Detergentes. Disolventes organicos: eter, cloroformo. Luz ultravioleta. Podofilinas, las cuales destruyen ademas tejidos del huesped. Farmacos Antivirales: farmacos que inhiben procesos especificos del virus, como la union a la celula, la decapsidacion del genoma viral o el ensamblaje de los nuevos virus, o, preferentemente, inhiben la sintesis macromolecular dirigida por el virus. Farmacos Inmunomodulares: comprenden los que reemplazan las respuestas inmunitarias deficientes del huesped y los que favorecen a los endogenos.
ANTIVIRALES QUE IMPIDEN LA ENTRADA DEL VIRUS A LA CELULA
Amantadina: actua sobre el virus influenza A. Su efecto principal es inhibir la decapsidacion viral o el desensamblaje del virion durante la endocitosis. Rimantadina: derivado 4-10 veces mas activa que la amantadina. Oseltamivir: inhibe la actividad de neuraminidasa del virus influenza. Zanamivir:inhibe la actividad de neuraminidasa del virus influenza. Docosanol: interfiere en la fusion del VHS-1 y VHS-2 a las membranas de las celulas del huesped. Es de uso topico.
ANTIVIRALES QUE ACTUAN SOBRE LA FASE DE REPLICACIÓN DEL GENOMA VIRAL
Aciclovir: inhibe la sintesis del ADN viral. Tiene accion sobre los herpesvirus. Valaciclovir: profarmaco del Aciclovir. Ganciclovir: inhibe la sintesis del ADN viral. Tiene accion sobre los herpesvirus, pero su caracteristica singular es una potente inhibicion de la replicacion del CMV. Valganciclovir: profarmaco del Ganciclovir. Adefovir: Actua como inhibidor competitivo de las polimerasas del ADN y transcriptasas inversas virales con respecto al dATP. Se selectividad se relaciona con una mayor afinidad por la ADN polimerasa del VHB que por las polimerasas de las celulas huesped. Esta aprobado para el tratamiento de la hepatitis B cronica. Cidofovir: inhibe la sintesis de ADN viral de los herpesvirus. Penciclovir: inhibe la sintesis de ADN viral de los herpesvirus. Famciclovir. Profarmaco del Penciclovir de administracion oral. Fomivirseno. Inhibe la replicacion del CMV humano. Se administra de forma intravitrea. Foscarnet: actua sobre los herpesvirus y la transcriptasa reversa del HIV. Es inhibidor frente a la mayoria de las cepas de CMV resistentes a ganciclovir y de VHS y VVZ resistentes a aciclovir. Idoxuridina. Inhibe la sintesis de ADN viral. Actua en especial sobre los herpesvirus y poxvirus. Ribavirina: se utiliza en el tratamiento de la infección por HCV y por el VSR. Trifluridina. Vidarabina
RETROVIRALES Retroviridae es una familia de virus que comprende los retrovirus. Son virus con envoltura que presentan un genoma de ARN monocatenario de polaridad + y se replican de manera inusual a través de una forma intermedia de ADN bicatenario. Este proceso se lleva a cabo mediante una enzima: la retrotranscriptasa o transcriptasa inversa, que dirige la síntesis de ADN a través de ARN y posee una importancia extraordinaria en la manipulación genética. Una vez se ha pasado de ARN monocatenario a ADN se inserta dentro del ADN propio de la célula infectada donde se comporta como un gen más (véase Ciclo reproductivo de los virus).1 Por tanto, se incluyen en el Grupo VI de la Clasificación de Baltimore.
2011-0175 Jafre Puello diapositiva # 1 virologia Virología es el estudio de virus y virus como agentes: clasificación y evolución, su estructura, sus formas de infectar y explotar las células para la reproducción del virus, las enfermedades que causan, las técnicas para aislar y la cultura de ellos y su uso en la investigación y terapia.
Virología es considerada una parte de la microbiología o de patología.
Una importante rama de la Virología es la clasificación de virus. Virus pueden clasificarse según la célula huésped que infectan: virus animales, virus de plantas, hongos virus y bacteriófagos (virus que infectan bacterias, que son los virus más complejos).
Otra clasificación utiliza la forma geométrica de su cápside (a menudo una hélice o un icosaedro) o la estructura del virus (por ejemplo, la presencia o la ausencia de un envolvente de lípidos). Gama de virus en tamaño de unos 30 nm a aproximadamente 450 nm, lo que significa que la mayoría de ellos no puede verse con microscopios de luces.
La forma y la estructura del virus ha sido estudiado por cristalografía de rayos X, microscopía electrónica y RMN.
El sistema de clasificación más útil y más ampliamente utilizado distingue virus segun el tipo de ácido nucleico que utilizan como material genético y el método de replicación viral que emplean coaxial células huésped en la producción de más virus:
Virus de ADN (divididas en virus de ADN bicatenario y los virus de ADN monocatenario mucho menos común), Virus ARN (dividieron en virus de ARN monocatenario de sentido positivo, virus de ARN monocatenario de sentido negativo y los virus de ARN bicatenario mucho menos común), invertir transcribir virus (virus de ADN de transcripción inversa de bicatenario y monocatenario transcripción inversa RNA virus incluyendo retrovirus).
La virología (rama de la microbiologia) es el estudio de los virus: su estructura, clasificación y evolución, su manera de infectar y aprovecharse de las células huésped para la reproducción del virus, su interacción con los organismos huéspedes, su inmunidad, la enfermedad que causan, las técnicas para su aislamiento, cultivo y su uso en investigación y terapia. La virología es considerada un sub-campo de la microbiología y la medicina. La virologia también es una rama taxonómica de la biología para estudiar los virus y todos sus componentes.
Su investigación incluye:
la replicación viral los patógenos virales. la inmunología viral. las vacunas virales. los métodos de diagnóstico. la quimioterapia antiviral. las medidas de control de una infección. los diferentes signos que manifiestan la presencia de virus.
Los virus presentan una amplia diversidad de formas y tamaños, llamadas «morfologías». Son unas 100 veces más pequeños que las bacterias. La mayoría de los virus estudiados tienen un diámetro de entre 10 y 300 nanómetros. Algunos Filovirus tienen un tamaño total de hasta 1.400 nm, sin embargo, sólo miden unos 80 nm de diámetro.71 La mayoría de virus no pueden ser observados con un microscopio óptico, de manera que se utilizan microscopios electrónicos de barrido y de transmisión para visualizar partículas víricas.72 Para aumentar el contraste entre los virus y el trasfondo se utilizan tinciones densas en electrones. Son soluciones de sales de metales pesados como wolframio, que dispersan electrones en las regiones cubiertas por la tinción. Cuando las partículas víricas están cubiertas por la tinción (tinción positiva), oscurecen los detalles finos. La tinción negativa evita este problema, tiñendo únicamente el trasfondo.73
Una partícula vírica completa, conocida como virión, consiste en un ácido nucleico rodeado por una capa de protección proteica llamada cápside. Las cápsides están compuestas de subunidades proteicas idénticas llamadas capsómeros.74 Los virus tienen un «envoltorio lipídico» derivado de la membrana celular del huésped. La cápside está formada por proteínas codificadas por el genoma vírico, y su forma es la base de la distinción morfológica.75 76 Las subunidades proteicas codificadas por los virus se autoensamblan para formar una cápside, generalmente necesitando la presencia del genoma viral. Sin embargo, los virus complejos codifican proteínas que contribuyen a la construcción de su cápside. Las proteínas asociadas con los ácidos nucleicos son conocidas como nucleoproteínas, y la asociación de proteínas de la cápside vírica con ácidos nucleicos víricos recibe el nombre de nucleocápside.77 78 En general, hay cuatro tipos principales de morfología vírica:
Helicoidal las cápsides helicoidales se componen de un único tipo de capsómero apilado alrededor de un eje central para formar una estructura helicoidal que puede tener una cavidad central o un tubo hueco. Esta formación produce viriones en forma de barra o de hilo, pueden ser cortos y muy rígidos, o largos y muy flexibles. El material genético, normalmente ARN monocatenario, pero a veces ADN monocatenario, queda unido a la hélice proteica por interacciones entre el ácido nucleico con carga negativa y la carga positiva de las proteínas. En general, la longitud de una cápside helicoidal está en relación con la longitud del ácido nucleico que contiene, y el diámetro depende del tamaño y la distribución de los capsómeros. El conocido virus del mosaico del tabaco es un ejemplo de virus helicoidal.79
Icosaédrica la mayoría de virus que infectan los animales son icosaédricos o casi-esféricos con simetría icosaédrica. Un icosaedro regular es la mejor manera de formar una carcasa cerrada a partir de subunidades idénticas. El número mínimo requerido de capsómeros idénticos es doce, cada uno compuesto de cinco subunidades idénticas. Muchos virus, como los rotavirus, tienen más de doce capsómeros y parecen esféricos, manteniendo esta simetría. Los ápices de los capsómeros están rodeados por otros cinco capsómeros y reciben el nombre de pentones. Las caras triangulares de éstos también se componen de otros seis capsómeros y reciben el nombre de hexones.
Envoltura algunas especies de virus se envuelven en una forma modificada de una de las membranas celulares, o bien es la membrana externa que rodea una célula huésped infectada, o bien membranas internas como la membrana nuclear o el retículo endoplasmático, consiguiendo así una bicapa lipídica exterior conocida como envoltorio vírico. Esta membrana se rellena de proteínas codificadas por el genoma vírico y el del huésped, la membrana lipídica en sí y todos los carbohidratos presentes son codificados completamente por el huésped. El virus de la gripe y el VIH utilizan esta estrategia. La mayoría de virus envueltos dependen de la envoltura para infectar.81
Es un taxón que afecta a vertebrados. Aden-(var.”adeno-“) Elemento prefijo del gr. “adén”, que significa “glándula”. Viriones isométricos no envueltos. Nucleocapside angular 70-90 nm diámetro. Simetría icosaedrica.
II- Virus de la peste porcina africana
Infecta a cerdos domesicos, facoceros, potamóquero de río y garrapatas. Causa fiebres hemorrágicas.
La peste africana se limitaba al continente que lleva su nombre hasta 1957 cuando la enfermedad se informó en otros países.
III- Arenaviridae
Virus que afecta a vertebrados.
Arena, viene del latín con la familia de términos del árido.
Estos virus son pequños, con envoltura y con forma icosaédrica.
V- Astroviridae
Virus que afecta a vertebrados.
Astro- es un prefijo lo cual significa estrellas.
Human astrovirus: Virones no envueltos. Nucleocapside isométrica. 27-30 nm diámetro. Simetría polihedrica. Con vértices.
VI- Baculoviridae
Virus que infecta vertebrados. Báculo: del latín, “palo”. Refiriéndose a la forma nucleocápside.
Son visibles en secciones de tejido infectado. Viriones ocluidos por cristales de proteínas o no ocluidos. Virion envuelto y pleomorfo forma de vara. Nucleocapsides en forma de vara. 200-450 nm long. Y 30-100 de diámetro.
VII- Badnavirus
Afecta a plantas
Etimología: Baciliforme Ba, DNA.
VIII- Barnaviridae
Infecta a hongos. Etimología: Baciliforme Ba, RNA. Sin envoltura, con un baciliformes T=1 simetría icosaédrica, aproximadamente 50 nm de longitud y 20 nm de gran tamaño.
IX- Birnaviridae
Afecta a vertebrados. Birna: para el ARN bipartito. No envueltos, de 60 nm de diámetro, virión icosaédricO con T=13 simetría.
X- Bromoviridae
Afecta a plantas. Bromo- porque infecta a Bromus inermis.
Viriones no envueltos, icosaédrico o baciliformes 26-35 nm de diámetro.
XI- Bunyaviridae
Afecta a vertebrados y a plantas. Viriones pleomorficos, esferoidales. “Bunyamwera” ciudad de Uganda donde se aisló.
Envueltos, esféricos. Diámetro de 80 a 120 nm.
XII- Caliciviridae
Virus que infecta a vertebrados y a invertebrados. “Calici” porque presenta depresiones calciformes.
Viriones no están envueltos y la capside es isométrica. Simetría icosaedrica.
XIII- Capillovirus
Infecta a plantas. Del latín “capella”, diminutivo de capa. El 3 es poliadenilado. Codifica por lo menos 3 proteínas.
Los virus son organismos tan pequeños, que su tamaño les permite pasar a través de los filtros que retienen bacterias. Las características básicas para identificar a un virus son: la composición del ácido nucleico, la disposición de la cápside, la presencia o ausencia de una envoltura, el tamaño del virus y el huésped.
MUESTRAS PARA DIAGNOSTICO:
Las muestras deben tomarse precozmente en la fase aguda de la infección. Deben transladarse al laboratorio a la mayor brevedad posible, en medio de transporte. Las muestras más comunes son:
• Torundas nasofaríngeas: adecuados para el diagnóstico de enterovirus, adenovirus, y herpes simplex. Las nasofaringeas se prefieren para el virus respiratorio sincicial y la mayoria de los otros respiratorios. Las nasales son las mejores para los rinovirus.
• Aspirados faríngeos: son mejores que los hisopos (torundas) nasofaringeos, pero estos últimos son mas prácticos.
• Torundas rectales y muestras de heces: Aquí se muestran virus no cultivables como rotavirus, o adenovirus, que se detectan por microscopía electrónica o con técnicas de detección de antígenos.
• Secreciones, Sangre o Líquido cefalorraquídeo.
• En algunas virosis excepcionalmente el diagnóstico solo puede efectuarse en Biopsia de cerebro, como en la encefalitis por herpes simplex; en Huéspedes inmunodeprimidos y con citomegalovirus, el virus solo puede identificarse en el órgano afectado.
Al carecer de las enzimas biosintéticas necesarias para su replicación, los virus no pueden replicarse en medios de cultivo inertes sino que necesitan de células vivas. La replicación se realiza en el interior de una célula huésped susceptible.
En los comienzos de la virología, se utilizaron embriones de pollo o pato y/o animales de experimentación para replicar los virus. Posteriormente, con el advenimiento de las técnicas para mantener vivas células “in vitro” se pudieron desarrollar numerosos cultivos celulares, aptos para la replicación de la mayoría de los virus que causan patología humana y animal. En la actualidad, para el aislamiento de virus se utilizan casi exclusivamente los cultivos celulares.
Luego de la penetración del virus en una célula susceptible, la replicación se lleva a cabo utilizando los mecanismos biosintéticos de la célula, dirigidos por la información contenida en el ácido nucleico viral. Este actúa como codificador de los mecanismos enzimáticos de síntesis de ácidos nucleicos y proteínas. De esta forma, la célula parasitada por un virus e vez de producir sus propios ácidos nucleicos y proteínas, recibe órdenes codificadas en el genoma viral parta sintetizar productos virales que, luego de su ensamble, darán lugar a la formación de nuevos viriones. Estos saldrán de la célula donde replicaron para iniciar nuevos ciclos de replicación en otras células susceptibles.
La replicación viral puede dividirse en las siguientes fases:
Adsorción: que depende de la interacción de proteínas de superficie del virus con receptores celulares específicos. Por ejemplo el receptor CD4 del linfocito T y la glicoproteína gp120 del virus HIV, las hemaglutininas de los virus influenza y los receptores de ácido acetil-neuramínico en la superficie de las células epiteliales, etc. Penetración y decapsidación: la penetración a través de la membrana plasmática de la célula puede efectuarse por los siguientes mecanismos: por pasaje directo, por fusión de la membrana celular con la envoltura viral, o por endocitosis o viropexis, mecanismo más frecuente, es un fenómeno similar a la fagocitosis, en el cual el virus es incluído en una vacuola fagocítica. Luego de la penetración se produce la decapsidación, es decir la liberación del ácido nucleico viral de la nucleocápside. Eclipse: en esta etapa no pueden observarse viriones en el interior de la célula, ni se recupera virus infeccioso. Latencia: este período va desde que se pierde la infectividad inicial hasta que se la recupera debido a la aparición de los nuevos viriones. Durante las etapas de eclipse y latencia ocurre la biosíntesis de los ácidos nucleicos y de las proteínas virales. Maduración: de produce el ensamble de los ácidos nucleicos neoformados con las proteínas de la cápside. Liberación: los nuevos viriones salen de la célula por lisis de la misma, como el virus polio; o bien por brotación a través de la membrana celular, como los paramixovirus o arenavirus; o de la membrana nuclear, como los herpesvirus o del retículo endoplásmico, como los togavirus
La adhesión es la propiedad de la materia por la cual se unen y plasman dos superficies de sustancias iguales o diferentes cuando entran en contacto, y se mantienen juntas por fuerzas intermoleculares.
La adhesión ha jugado un papel muy importante en muchos aspectos de las técnicas de construcción tradicionales. La adhesión del ladrillo con el mortero (cemento) es un ejemplo claro.
Mecanismos de adhesión:
La cohesión es la causa de que el agua forme gotas, la tensión superficial hace que se mantengan esférica y la adhesión la mantiene en su sitio. Las gotas de agua son más planas sobre la flor de Hibiscus ya que tienen mejor adhesión.
Cinco mecanismos han sido propuestos para explicar por qué un material se adhiere a otro. Adhesión mecánica
Los materiales adhesivos rellenan los huecos o poros de las superficies manteniendo las superficies unidas por enclavamiento. Existen formas a gran escala de costura, otras veces a media escala como el velcro y algunos adhesivos textiles que funcionan a escalas pequeñas. Es un método similar a la tensión superficial Adhesión química
Dos materiales pueden formar un compuesto químico al unirse. Las uniones más fuertes se producen entre átomos donde hay permutación (enlace iónico) o se comparten electrones (enlace covalente). Un enlace más débil se produce cuando un átomo de hidrógeno que ya forma parte de una partícula se ve atraída por otra de nitrógeno, oxígeno o flúor, en ese caso hablaríamos de un puente de hidrógeno. Adhesión dispersiva
En la adhesión dispersiva, dos materiales se mantienen unidos por las fuerzas de van der Waals: la atracción entre dos moléculas, cada una de las cuales tiene regiones de carga positiva y negativa. En este caso, cada molécula tiene una región de mayor carga positiva o negativa que se une a la siguiente de carga contraria. Este efecto puede ser una propiedad permanente o temporal debido al movimiento continuo de los electrones en una región.
En la ciencia de superficies el término "adhesión" siempre se refiere a una adhesión dispersiva. En un sistema sólido-líquido-gas normal (como una gota de un líquido sobre una superficie rodeada de aire) el ángulo de contacto es usado para cuantificar la adhesividad. En los casos donde el ángulo de contacto es bajo la adhesión está muy presente. Esto se debe a que una mayor superficie entre el líquido y el sólido conlleva una energía superficial mayor. Adhesión electrostática
Algunos materiales conductores dejan pasar electrones formando una diferencia de potencial al unirse. Esto da como resultado una estructura similar a un condensador y crea una fuerza electrostática atractiva entre materiales. mi con diversos materiales les pueden ayudar
Penetración y decapsidación: la penetración a través de la membrana plasmática de la célula puede efectuarse por los siguientes mecanismos: por pasaje directo, por fusión de la membrana celular con la envoltura viral, o por endocitosis o viropexis, mecanismo más frecuente, es un fenómeno similar a la fagocitosis, en el cual el virus es incluído en una vacuola fagocítica. Luego de la penetración se produce la decapsidación, es decir la liberación del ácido nucleico viral de la nucleocápside.
Penetración de los Bacteriófagos:
Inyección del material genético viral: Después de la adsorción, se produce un cambio configuracional en las proteínas de la placa basal, alguna de las cuales tienen actividad enzimática y producen un poro en la membrana citoplasmática de la célula. La vaina del fago se contrae y el material genético viral ingresa en la célula, mientras que la envoltura proteica queda en el exterior. Para entrar en una célula, los fagos se acoplan a receptores específicos en la superficie de la bacteria, que pueden ser lipopolisacáridos, ácidos teicoicos, proteínas o incluso flagelos. Por ello, cada fago solo podrá infectar ciertas bacterias según sus receptores. Puesto que los fagos no son móviles, dependen de encuentros al azar con los receptores adecuados en solución para poder infectar una bacteria. Parece que los bacteriófagos presentan una especie de jeringa mediante la cual introducen su material genético en el interior de la célula. Tras el reconocimiento del receptor adecuado, la cola y cuello del fago se contraen, quedando así el fago acoplado a la superficie celular. El material genético puede ser ahora introducido a través de la membrana o bien simplemente depositado sobre la superficie. No se descarta que pueda haber fagos con otros métodos diferentes para introducir su material genético en la célula.
partes 3 y 4 dios te bendiga 2011-0119 LA CÉLULA BACTERIANA Las bacterias son células muy sencillas; carecen de núcleo y tampoco presentan orgánulos en el citoplasma. Se las denomina Procariotas. Son organismos unicelulares y se encuentran en todos los ecosistemas. Las bacterias son un numeroso grupo de seres vivos, con características muy diversas. En la clasificación de los Dominios, Woese, aparecen dos grupos de Procariotas: el Dominio Archaea, que engloba a los organismos más antiguos del Planeta, y el Dominio Bacteria, en el que se encuentran la gran mayoría de los organismos bacterianos actuales, también conocidos con el nombre de Eubacterias. Las bacterias se caracterizan por no tener núcleo patente, ni clorofila típica, aunque pueden poseer otros pigmentos similares como bacterioclorina, bacterioclorofila y bacteriopurpurina, y sin pseudópodos. La bacterioclorina es un pigmento verde fotosintetizante que, según ciertos autores, acompaña a la bacteriopurpurina y a la bacterioclorifila en las algas purpúreas.
Tamaño de los Virus
Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus.
Su pequeño tamaño explica lo tardío del descubrimiento de estos agentes. La primera referencia sobre la existencia de los virus se debe al botánico ruso Dimitri Ivanovski en 1892. Este investigador buscaba el agente causante de la enfermedad denominada mosaico del tabaco, y llegó a la conclusión de que debía tratarse de una toxina o de un organismo más pequeño que las bacterias, pues el agente atravesaba los filtros que retenían las bacterias. Denominó a estos agentes patógenos virus filtrables.
La virología (rama de la microbiologia) es el estudio de los virus: su estructura, clasificación y evolución, su manera de infectar y aprovecharse de las células huésped para la reproducción del virus, su interacción con los organismos huéspedes, su inmunidad, la enfermedad que causan, las técnicas para su aislamiento, cultivo y su uso en investigación y terapia. La virología es considerada un sub-campo de la microbiología y la medicina. La virologia también es una rama taxonómica de la biología para estudiar los virus y todos sus componentes.
Su investigación incluye: la replicación viral los patógenos virales. la inmunología viral. las vacunas virales. los métodos de diagnóstico. la quimioterapia antiviral. las medidas de control de una infección. los diferentes signos que manifiestan la presencia de virus.
Un virus está compuesto de una molécula de ácido nucleico y una envoltura proteínica. Ésta es la estructura básica de un virus, aunque algunos de ellos pueden añadir a esto la presencia de alguna enzima, bien junto al ácido nucleico, como la transcriptasa inversa de los retrovirus, bien en la envoltura, para facilitar la apertura de una brecha en la membrana de la célula hospedadora.
A la unidad formada por el ácido nucleico y la envoltura proteínica se le denomina tambiénvirión.
El ácido nucleico es solamente de un tipo, ADN o ARN, nunca los dos.
Atendiendo al tipo de ácido nucleico se distinguen cuatro clases de virus:
- ADN de cadena doble
- ADN de cadena sencilla
- ARN de cadena doble
- ARN de cadena sencilla
La envoltura proteínica recibe el nombre de cápsida. Está formada por unas subunidades idénticas denominadas capsómeros. Los capsómeros son proteínas globulares que en ocasiones tienen una parte glicídica unida. Se ensamblan entre sí dando a la cubierta una forma geométrica.
Atendiendo la forma de la cápsida, se pueden distinguir los siguientes tipos de virus:
- Cilíndricos o helicoidales: los capsómeros, que son de un solo tipo, se ajustan entorno una hélice simple de ácido nucleico. Un ejemplo lo constituye el virus del mosaico del tabaco.
- Icosaédricos: los capsómeros, que suelen ser de varios tipos, se ajustan formando un icosaedro regular (es decir, 20 caras triangulares y 12 vértices), y dejando un hueco central donde se sitúa el ácido nucleico fuertemente apelotonado. Algunos forman poliedros con más caras que el icosaedro, y algunos presentan fibras proteicas que sobresalen de la cápsida.
Un ejemplo lo constituyen los adenovirus, entre los que se encuentran los virus de los resfriados y faringitis.
La simetría es la forma que adopta un virus en el espacio y está dad por la estructura de su nucleocápside. La simetría puede ser helicoidal, icosaédrica o compleja.
Los virus con simetría helicoidal presentan una nucleocápside cilindrica que puede estar extendida como en el virus del mosaico del tabaco. Este es un virus desnudo. En otros casos la nucleocápside está arrollada sobre sí misma y recubierta por una envoltura, como en el virusInfluenza.
Los virus con simetría icosaédrica son poliedros regulares con 20 caras triangulares, 30 aristas y 12 vértices. Son ejemplos de virus con simetría icosaédrica desnudos el virus de la poliomielitis o losadenovirus. Los herpesvirus y togavirus son virus con simetría icosaédrica y recubiertos por una envoltura.
Se denominan virus de simetría compleja a aquellos que presentan una nucleocápside helicoidal o icosaédrica recubierta por una envoltura laxa. Ejemplos de éstos virus son los poxvirus (forma de ladrillo), los rhabdovirus (forma de bala) y algunos bacteriófagos que poseen una cabeza con simetría icosaédrica y una cola con simetría helicoidal.
Simetría cúbica: Toda la simetría cúbica observada en los virus de los animales, hasta la fecha, es del patrón icosaédrico, la disposición más eficiente de subunidades en una cubierta cerrada. Conocer las reglas que orientan la simetría icosaédrica hace posible determinar el número de capsómeros en una partícula, característica importante de la clasificación de los virus.
El ácido nucleico viral se condensa dentro de las partículas isoméricas; en la condensación del ácido nucleico en una forma adecuada para su acumulación participan proteínas "nucleares" codificadas por el virus o, en el caso de los papovirus, histonas celulares. No han podido definirse aún las reglas para la incorporación del ácido nucleico en las partículas isométricas; probablemente una "secuencia de empaque" participa en el ensamble, aunque en general no son cruciales las estructuras primarias, secundarias y terciarias del ácido nucleico. Existen limitaciones de tamaño en las moléculas de ácido nucleico que se pueden empacar o acumular en una cápside icosaédrica determinada. Las cápsides icosaédricas se forman de manera independiente del ácido nucleico. La mayor parte de los preparados de virus isométricos contendrán algunas partículas "vacías", desprovistas de ácido nucleico viral. Ejemplos de simetría cúbica son los grupos de virus tanto de ADN como de ARN.
Simetría helicoidal: En los casos de simetría helicoidal, las subunidades proteínicas están fijas de manera periódica al ácido nucleico viral, con lo que lo amplían en una espiral. El complejo filamentoso de proteína y ácido nucleico viral (nucleocápside) se enrolla a continuación dentro de una cubierta que contiene lípidos. Por lo tanto, a diferencia del caso de las estructuras icosaédricas, existe una interacción periódica regular entre la proteína de la cápside y el ácido nucleico en el virus que tiene simetría helicoidal. No es posible que se formen partículas helicoidales "vacías".
Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus.
Su pequeño tamaño explica lo tardío del descubrimiento de estos agentes. La primera referencia sobre la existencia de los virus se debe al botánico ruso Dimitri Ivanovski en 1892. Este investigador buscaba el agente causante de la enfermedad denominada mosaico del tabaco, y llegó a la conclusión de que debía tratarse de una toxina o de un organismo más pequeño que las bacterias, pues el agente atravesaba los filtros que retenían las bacterias. Denominó a estos agentes patógenos virus filtrables.
En 1897, el microbiólogo holandés Martinus Beijerink realizó experimentos similares a los de Ivanovski, y llegó a desechar la idea de las toxinas, pues se trataba de un agente capaz de reproducirse, ya que mantenía su poder infeccioso de unas plantas a otras, sin diluirse su poder patógeno. Poco después, los microbiólogos alemanes Frederick Loeffler y Paul Frosch descubrieron que la fiebre aftosa del ganado era producida por un virus filtrable que actuaba como un agente infeccioso.
En la década de los 30, con el uso de filtros de tamaño de poro inferior, con las técnicas de cultivo celular in vitro que permitían la obtención de gran cantidad de virus, con la ultracentrifugación y finalmente con el microscopio electrónico y la difracción de rayos X, se logró visualizar a estos agentes.
Los virus son cristalizables, como demostró W. Stanley en 1935. Esto depende del hecho de que las partículas víricas tienen formas geométricas precisas y que son idénticas entre sí, lo cual las separa de la irregularidad característica de los organismos, las células o los orgánulos, y las acerca a las características de los minerales y de agregados de macromoléculas como los ribosomas. Al tener un volumen y forma idénticos, las partículas víricas tienden a ordenarse en una pauta tridimensional regular, periódica, es decir, tienden a cristalizar.
1- Picornaviridae: es una familia de virus infectivos para animales. Contienen un genoma ARN monocatenario positivo y por lo tanto se incluyen en el Grupo IV de la Clasificación de Baltimore. El genoma ARN es inusual porque tiene una proteína en el terminal 5' que se utiliza como iniciador de la transcripción por la ARN polimerasa. El nombre se deriva de "pico", que significa pequeño, con lo que "picornavirus" significa literalmente "virus RNA pequeños".
2- Caliciviridae: es una familia de virus infectivos para animales y causantes de gastroenteritis en humanos. Los calicivirus han sido encontrados en la mayoría de los animales domésticos y muchos silvestres, como cerdos, conejos, gallinas y anfibios. Contienen un genoma ARN monocatenario positivo y por lo tanto se incluyen en el Grupo IV de la Clasificación de Baltimore.
3- Hepadnaviridae es una familia de virus que causan infecciones en hígado de humanos y de animales. Los hepadnavirus tienen un genoma muy corto de ADN parcialmente de doble hélice y parcialmente de hélice simple, circular. Como su replicación involucra un ARN intermedio, se incluyen en el Grupo VII de la Clasificación de Baltimore.
4- Reoviridae es una familia de virus que pueden afectar el sistema gastrointestinal (tales como rotavirus ) y tracto respiratorio . Los virus de la familia Reoviridae han genomas que consiste en segmentar, ARN de doble cadena (dsRNA). El nombre "Reoviridae" se deriva de las vías respiratorias entéricas virus huérfanos.
5- Rhabdoviridae es una familia de virus infectivos para animales y plantas. Entre los animales infectados están insectos, peces y mamíferos, incluidos los humanos. Estos virus presentan una forma alargada parecida a una bala. Poseen un genoma de ARN monocatenario de sentido negativo por lo que se incluye en el Grupo V de la Clasificación de Baltimore.
6- El Togaviridae es una familia de virus que incluye los siguientes géneros: Género Alphavirus; especies típicas: Sindbis virus, Virus de la encefalitis equina oriental, Virus de la encefalitis equina occidental, Virus de la encefalitis equina venezolana, Virus del Río Ross, Virus O'nyong'nyong. Género Rubivirus; especies típicas: Rubéola virus.
7- Los Coronavirus son un género de virus ARN de vertebrados de Coronaviridae.1 Son virus envueltos con un genoma de ARN de cadena sencilla con polaridad positiva y simetría helicoidal. El tamaño genómico de los coronavirus varía entre 16 a 31 kilobases, un virus de extraordinaria longitud.
8- Bunyavirus, género de virus ARN de vertebrados de la familia Bunyaviridae.
9- Papovaviridae es una familia de virus pequeños (apróx. 50 nm) infectivos para una gran variedad de animales, incluyendo los humanos. Su nombre implica las patología que causa: papilomavirus +
10- Los paramixovirus son miembros de la familia de virus paramyxoviridae, que poseen un genoma con ARN de cadena sencilla de polaridad negativa como ácido nucleico; albergan dicha información genética en una cápside cubierta de envoltura viral y estructuralmente definida por una simetría helicoidal, de un tamaño de 125 a 250 x 18 nm, con envoltura, pleomórfico, y un filamentoso nucleocápside; y por ensamblar los viriones maduros en el citoplasma como compartimento celular 11- Los Orthomyxoviridae son una familia de virus RNA que infectan a los vertebrados. Incluyen a los virus causantes de la gripe.
12- Un retrovirus es un virus de ARN que se replica en una célula huésped a través del proceso de transcripción inversa . En primer lugar, utiliza su propio transcriptasa inversa enzima para producir ADN de su ARN del genoma, la inversa de la pauta habitual, por lo tanto retro (hacia atrás).
13- Flavivirus : es un género de virus ARN pertenecientes a la familia Flaviviridae. Los Flavivirus son virus con envoltura, la simetría de la nucleocápside icosaedrica, y cuyo material genético reside en una única cadena de ARN de polaridad positiva.
14- Parvovirus, a veces truncada a "parvo", es a la vez el nombre común en Inglés casualmente se aplica a todos los virus de la Parvoviridae familia taxonómica y también el nombre taxonómico del parvovirus género dentro de la Parvoviridae familia.
15- Los adenovirus (Adenoviridae) son una familia de virus que infectan tanto humanos como animales. Son virus no encapsulados de ADN bicatenario que pueden provocar infecciones en las vías respiratorias, conjuntivitis, cistitis hemorrágica y gastroenteritis. Los adenovirus también se utilizan para obtener ADN para la terapia génica.
16- El Arenavirus es un género de virus de la familia Arenaviridae.La especie tipo es el virus de la coriomeningitis linfocítica (LCMV); es también de las spp. responsables de la fiebre Lassa. Arena, viene del latín con la familia de términos del árido.
17- Los herpesvirus o virus de la familia Herpesviridae, deben su nombre al término griego herpein (ἕρπειν), reptar o arrastrar, haciendo alusión a la facultad de estos agentes infecciosos microscópicos de ser fácilmente contagiados y transmitidos de una persona a la otra y de recurrencia crónica. Los herpesvirus forman una familia de virus divididos en tres subfamilias: alpha, beta y gammaherpesvirinae, ubicados con base en la arquitectura del virión y ciertas propiedades biológicas comunes.
18- Poxviridae es una familia de virus de ADN relacionados entre sí llamados poxvirus, infectivos para animales vertebrados (8 de los 11 géneros, por ejemplo, poxvirus de la gallina) y a los invertebrados (3 géneros). Tienen un genoma ADN bicatenario y por lo tanto pertenece al Grupo I de la Clasificación de Baltimore.
19- Filoviridae es una familia de virus del orden Mononegavirales. Se caracterizan por poseer como genoma RNA monocatenario de sentido negativo, y por infectar a primates. Agrupa, entre otros, al virus Ébola y al virus Marburg.
Los virus son organismos tan pequeños, que su tamaño les permite pasar a través de los filtros que retienen bacterias. Las características básicas para identificar a un virus son: la composición del ácido nucleico, la disposición de la cápside, la presencia o ausencia de una envoltura, el tamaño del virus y el huésped.
MUESTRAS PARA DIAGNOSTICO:
Las muestras deben tomarse precozmente en la fase aguda de la infección. Deben transladarse al laboratorio a la mayor brevedad posible, en medio de transporte. Las muestras más comunes son:
• Torundas nasofaríngeas: adecuados para el diagnóstico de enterovirus, adenovirus, y herpes simplex. Las nasofaringeas se prefieren para el virus respiratorio sincicial y la mayoria de los otros respiratorios. Las nasales son las mejores para los rinovirus.
• Aspirados faríngeos: son mejores que los hisopos (torundas) nasofaringeos, pero estos últimos son mas prácticos.
• Torundas rectales y muestras de heces: Aquí se muestran virus no cultivables como rotavirus, o adenovirus, que se detectan por microscopía electrónica o con técnicas de detección de antígenos.
• Secreciones, Sangre o Líquido cefalorraquídeo.
• En algunas virosis excepcionalmente el diagnóstico solo puede efectuarse en Biopsia de cerebro, como en la encefalitis por herpes simplex; en Huéspedes inmunodeprimidos y con citomegalovirus, el virus solo puede identificarse en el órgano afectado.
DIAGNOSTICO VIRAL
Observación del efecto citopático, en cultivo de tejidos inoculados con ciertos virus. La etiología de un determinado síndrome viral puede demostrarse recuperando el agente causal en tejidos celulares de diversos tipos (por ej: riñón de mono, pulmón humano fetal, amnios humano o células tumorales humanas). Los huevos embrionados actualmente solo se utilizan para cultivar el virus cuando se desea preparar vacunas. El empleo de animales de experimentación es excepcional en el diagnóstico.
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada. La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos. Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante.
Fijación, penetración y pérdida de la cubierta: La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado. Después de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original.
ciclo litico de un bacteriófago.formado por acido nuceico ADN y capsida compleja.fijacion o adsorcion: se fijan inicialmente a traves de las puntas de las fibras cudales mediante enlaces quimicos y posterormente de forma mecanica al clavar las pespinas basales en la pared bacteriana.
penetracion: el bacteriofago mediante enzimas lisozimas situadas en su placa basal perrfora la pared cecular y luego contrae la vaina e introduce el eje tubular de modo que el adn pasa directamente al citoplasma
Los virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación. Síntesis de los componentes virales: Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo). Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
La adhesión es la propiedad de la materia por la cual se unen y plasman dos superficies de sustancias iguales o diferentes cuando entran en contacto, y se mantienen juntas por fuerzas intermoleculares. La adhesión ha jugado un papel muy importante en muchos aspectos de las técnicas de construcción tradicionales. La adhesión del ladrillo con elmortero (cemento) es un ejemplo claro. La cohesión es distinta de la adhesión. La cohesión es la fuerza de atracción entre partículas adyacentes dentro de un mismo cuerpo, mientras que la adhesión es la interacción entre las superficies de distintos cuerpos.1 Mecanismos de adhesión Adhesión mecánica Los materiales adhesivos rellenan los huecos o poros de las superficies manteniendo las superficies unidas por enclavamiento. Existen formas a gran escala de costura, otras veces a media escala como el velcro y algunos adhesivos textiles que funcionan a escalas pequeñas. Es un método similar a la tensión superficial Adhesión química Dos materiales pueden formar un compuesto químico al unirse. Las uniones más fuertes se producen entre átomos donde hay permutación (enlace iónico) o se comparten electrones (enlace covalente). Un enlace más débil se produce cuando un átomo de hidrógeno que ya forma parte de una partícula se ve atraída por otra de nitrógeno, oxígeno o flúor, en ese caso hablaríamos de un puente de hidrógeno. Adhesión dispersiva En la adhesión dispersiva, dos materiales se mantienen unidos por las fuerzas de van der Waals: la atracción entre dos moléculas, cada una de las cuales tiene regiones de carga positiva y negativa. En este caso, cada molécula tiene una región de mayor carga positiva o negativa que se une a la siguiente de carga contraria. Este efecto puede ser una propiedad permanente o temporal debido al movimiento continuo de los electrones en una región.. Adhesión electrostática Algunos materiales conductores dejan pasar electrones formando una diferencia de potencial al unirse. Esto da como resultado una estructura similar a un condensador y crea una fuerza electrostática atractiva entre materiales. mi con diversos materiales les pueden ayudar
Los bacteriófagos (también llamados fagos -del griego φαγητόν (phagētón), «alimento, ingestión») son virus que infectan exclusivamente a lasbacterias. Al igual que los virus que infectan células eucariotas, los fagos están constituidos por una cubierta proteica o cápside en cuyo interior está contenido su material genético, que puede ser ADN o ARN de simple o doble cadena, circular o lineal (en el 95% de los fagos conocidos es ADN de doble cadena), de 5.000 a 500.000 pares de bases. El tamaño de los fagos oscila entre 20 y 200 nm aproximadamente. Los fagos son ubicuos y pueden ser encontrados en diversas poblaciones de bacterias, tanto en el suelo como en la flora intestinal de los animales. Uno de los ambientes más poblados por fagos y otros virus es el agua de mar, donde se estima que puede haber en torno a 109partículas virales por mililitro, pudiendo estar infectadas por fagos el 70% de las bacterias marinas.
Historia
La historia del descubrimiento de los bacteriofagos ha sido objeto de muchos debates, los cuales incluyen una controversia sobre quién fue su descubridor. En 1913, el bacteriólogo británicoFrederick Twort descubrió un agente bacteriolítico que infectaba y mataba a las bacterias, pero, no fue capaz de saber qué era exactamente dicho agente, señalando en una de sus hipótesis que, entre otras posibilidades, podría tratarse de un virus. No fue sino hasta 1917, cuando el microbiólogo canadiense Félix d'Herelle anunció el descubrimiento de "un invisible antagonista microbiano del bacilo de la disentería", al cual afirmó que se trataba de un virus al cual llamó bacteriófago. Los trabajos de d'Herelle iniciaron desde 1910 y en contraste con Twort y algunos otros científicos que habían reportado fenómenos similares, tuvo pocas dudas sobre la naturaleza del fenómeno que estaba observando y afirmó que se trataba de un virus que parasitaba a la bacteria. El nombre bacteriófago lo formó de la palabra "bacteria" y "phagein" (comer o devorar, en griego), implicando que los fagos "comen" o "devoran" a las bacterias.
continuacion partes 7 2011-0119 dios te bendiga Diagnostico de la Virosis
Los virus son organismos tan pequeños, que su tamaño les permite pasar a través de los filtros que retienen bacterias. Las características básicas para identificar a un virus son: la composición del ácido nucleico, la disposición de la cápside, la presencia o ausencia de una envoltura, el tamaño del virus y el huésped.
MUESTRAS PARA DIAGNOSTICO:
Las muestras deben tomarse precozmente en la fase aguda de la infección. Deben transladarse al laboratorio a la mayor brevedad posible, en medio de transporte. Las muestras más comunes son:
• Torundas nasofaríngeas: adecuados para el diagnóstico de enterovirus, adenovirus, y herpes simplex. Las nasofaringeas se prefieren para el virus respiratorio sincicial y la mayoria de los otros respiratorios. Las nasales son las mejores para los rinovirus.
• Aspirados faríngeos: son mejores que los hisopos (torundas) nasofaringeos, pero estos últimos son mas prácticos.
• Torundas rectales y muestras de heces: Aquí se muestran virus no cultivables como rotavirus, o adenovirus, que se detectan por microscopía electrónica o con técnicas de detección de antígenos.
Mat:2011-0289 Maduracion y salida de virus envueltos y desnudos.
Los virus cubiertos maduran por un proceso de gemación. Se insertan glucoproteínas de cubierta específica del virus en las membranas celulares y, a continuación, las nucleocápsides hacen gemación a través de la membrana a nivel de estos sitios modificados y, al hacerlo así, adquieren su cubierta. La gemación ocurre a menudo a nivel de la membrana plasmática y puede abarcar a otras membranas de la célula. Los virus cubiertos no son infecciosos hasta que han adquirido sus cubiertas. Por lo tanto, no se acumulan de manera típica viriones descendientes infecciosos en el interior de la célula infectada.
La maduración viral es a veces un proceso ineficaz. Pueden acumularse cantidades excesivas de componentes virales y participar en la formación de cuerpos de inclusión en la célula (ver antes). Como resultado de los profundos efectos dañinos de la replicación viral, se desarrollan por último efectos citopáticos celulares y la célula muere. Sin embargo, hay casos en que el virus no lesiona a la célula y se producen infecciones persistentes y prolongadas.os genomas virales recién sintetizados y los polipéptidos de la cápside se ensamblan para formar los virus descendientes. Las cápsides icosaédricas pueden condensarse en ausencia de ácido nucleico, en tanto que las nucleocápsides de los virus de simetría helicoidal no pueden formarse sin ARN viral.
No existen mecanismos especiales para la liberación de virus carentes de cubierta; las células infectadas acaban por experimentar lisis y liberar las partículas virales.
La adhesión es la propiedad de la materia por la cual se unen y plasman dos superficies de sustancias iguales o diferentes cuando entran en contacto, y se mantienen juntas por fuerzas intermoleculares.
Mecanismos de adhesion
La cohesión es la causa de que el agua forme gotas, la tensión superficial hace que se mantengan esférica y la adhesión la mantiene en su sitio.
Las gotas de agua son más planas sobre la flor de Hibiscus ya que tienen mejor adhesión.
Cinco mecanismos han sido propuestos para explicar por qué un material se adhiere a otro.
Adhesión mecánica
Los materiales adhesivos rellenan los huecos o poros de las superficies manteniendo las superficies unidas por enclavamiento. Existen formas a gran escala de costura, otras veces a media escala como el velcro y algunos adhesivos textiles que funcionan a escalas pequeñas. Es un método similar a la tensión superficial.
Adhesión química
Dos materiales pueden formar un compuesto químico al unirse. Las uniones más fuertes se producen entre átomos donde hay permutación (enlace iónico) o se comparten electrones (enlace covalente). Un enlace más débil se produce cuando un átomo de hidrógeno que ya forma parte de una partícula se ve atraída por otra de nitrógeno, oxígeno o flúor.
Penetración de los Bacteriófagos
Los virus son moléculas de DNA o RNA rodeadas por una envoltura proteica que necesitan células viables para poder replicarse. Los virus utilizan la maquinaria metabólica de las células para sintetizar su material genético y proteínas de la envoltura. Existen distintos tipos de virus que pueden infectar células procariontes o células eucariontes. Los bacteriófagos o fagos son virus que se reproducen en células procariontes. Replicacion:
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada
Los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes.
No todas las partículas víricas liberadas son infecciosas. La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
Los virus envueltos (como el VIH) son liberados de la célula huésped por gemación. Durante este proceso, el virus adquiere su envoltura, que es una parte modificada de la membrana plasmática del huésped. Maduración: de produce el ensamble de los ácidos nucleicos neoformados con las proteínas de la cápside. Liberación: los nuevos viriones salen de la célula por lisis de la misma, como el virus polio; o bien por brotación a través de la membrana celular, como los paramixovirus o arenavirus; o de la membrana nuclear, como los herpesvirus o del retículo endoplásmico, como los togavirus.
continuacion 2011-0119 dios te bendiga Maduración y ensamblaje
Cuando se han sintetizado un número crítico de varios componentes virales, éstos se ensamblan en partículas víricas maduras en el núcleo y/o citoplasma de la célula infectada. En este ensamblaje y al contrario de lo que ocurre en los bacteriófagos, no están implicados enzimas biosintéticos especiales, sino que ocurre de forma espontánea debido a la gran afinidad de la cápsida con el ácido nucleico viral. La maduración se puede definir como la fase de la infección viral durante la cual los componentes estructurales del virión se producen y ensamblan con el nuevo ácido nucleico viral para formar la nucleocápsida.
Salida de un Virus Envuelto
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula
Las drogas antirretrovirales actúan sobre diferentes pasos necesarios para la replicación viral; hasta la fecha, las más extendidas son aquellas drogas que actúan sobre la transcriptasa inversa y las drogas que actúan sobre las proteasas; aunque ya existen fármacos capaces de inhibir otros sitios tales como los inhibidores de la fusión y los de la integrasa. Los Inhibidores de la Reversotranscriptasa (IRT) alteran la función de la enzima llamada reversotranscriptasa que es la que utiliza el VIH para cambiar su mensaje genético a una forma que pueda ser fácilmente insertada dentro del núcleo de la célula infectada. Es la encargada de hacer una copia en ADN del ARN viral de manera que éste pueda insertarse en el ADN celular, mediante la acción de las integrasas, y así producir proteínas del VIH necesarias para la auto reproducción viral. Los IRT nucleósidos y los nucleótidos necesitan ser fosforilados para luego interactuar con un sustrato en el sitio de unión a la enzima y, cuando es incorporado, entonces termina la elongación de la cadena de ADN; mientras que los IRT no nucleósidos no necesitan ninguna conversión metabólica y bloquean directamente la reacción con la reverso transcriptasa por una interacción específica con el sitio de unión “sin sustrato” de la reversotranscriptasa del VIH. Los Inhibidores de las Proteasas (IP) son fármacos con una potente actividad antiviral. La proteasa del VIH es requerida para el clivaje del polipéptido gag-pol en sus partes funcionales y la inhibición de este clivaje trae como consecuencia una disminución en la producción de viriones maduros. O sea, estos medicamentos se parecen a los trozos de la cadena de proteína que la proteasa corta normalmente. Al adherirse a la proteasa, los IP previenen que la proteasa corte las cadenas largas de proteínas y enzimas en trozos más cortos que el virus necesita para reproducirse en nuevas copias de sí mismo. Aunque las cadenas largas no hayan sido cortadas en los trocitos correctos, las nuevas copias del VIH aún se construyen y continúan empujando a través de la membrana de la célula infectada. Pero estas nuevas copias del VIH son “defectuosas”, ya que no han sido formadas completamente, por lo tanto no pueden continuar infectando otras células.(1,2,3,4) INHIBIDORES DE FUSION Los inhibidores de la entrada evitan el ingreso del VIH a las células sanas, se adhieren a las proteínas de superficie de las células T o a las proteínas de superficie del VIH. Para que el VIH es una y pueda entrar a las células T, las proteínas de la capa externa del VIH deben unirse a las proteínas de superficie de las células T. Los inhibidores de la entrada evitan que este proceso ocurra. Algunos inhibidores de la entrada se adhieren a las proteínas gp120 ó gp41 de la capa externa del VIH. Otros inhibidores utilizan la proteína CD4 ó los receptores CCR5 ó CXCR4 de la superficie de las células T. Si los inhibidores de la entrada logran adherirse eficientemente a las proteínas, éstas quedan bloqueadas y entonces el VIH no puede unirse a la superficie de las células T e ingresar a las mismas.
MECANISMO DE ACCION DE LOS RETROVIRALES
Los antivirales son un tipo de medicamento usado para el tratamiento de infecciones producidas por virus. Tal como los antibióticos (específicos para bacteria), existen antivirales específicos para distintos tipos de virus. No sin excepciones, son relativamente inocuos para el huésped, por lo que su aplicación es relativamente segura. Deben distinguirse de los viricidas, que son compuestos químicos que destruyen las partículas virales presentes en el medio ambiente. Su eficacia (y a veces su toxicidad) están relacionadas con la competencia inmunológica del paciente. Su actividad es restringida al actuar sobre un “órgano diana” muy específico. Desarrollo de Resistencias. La mayoría solo son eficaces durante la fase de Replicación viral y no sobre los virus Latentes. La utilización óptima de los antivirales precisa de la realización de un diagnóstico clínico correcto.
El término acelular significa literalmente "sin células". Se aplica principalmente a los virus, pero también a provirus y viroides, que son entidades biológicas no formadas por células, a diferencia de bacterias, compuestas por células procariotas, y de hongos, vegetales y animales, formados por células eucariotas.
LOS VIROIDES I y II
son agentes infecciosos que, al igual que los virus, tienen un ciclo extracelular que se caracteriza por la inactividad metabólica y unciclo intracelular en el que causan infección al huésped susceptible, pero que a diferencia de los virus, los viroides no poseen proteínas ni lípidos y están constituidos por una cadena cíclica corta de ARN, circular o con forma de varilla, (que no codifica proteínas).1 Es importante decir que tanto su forma intracelular como extracelular son las mismas (ARN desnudo), los mecanismos por los cuales éstos logran causar infección están relacionados con la autocatálisis de su material genético. Estas pequeñas moléculas de ARN circular de cadena sencilla formadas por unos pocos cientos de nucleótidos. Carecen de cápsida y su tamaño es una milésima parte del de los virus más pequeños. - Aparecen con mayor abundancia en el núcleo de las células infectadas. - Sólo han sido detectados en plantas, en las que producen una gran variedad de enfermedades y pueden ser transmitidos tanto horizontal como verticalmente. - No existen ninguna evidencia de que los viroides son traducidos a proteínas, ni se conoce cómo causan las enfermedades. Lo que sí se sabe es que son replicados por los enzimas del huésped. - Algunas semejanzas entre la secuencia de nucleótidos de los viroides y ciertos secuencias del genoma de las plantas no traducidas (intrones) sugieren que los viroides evolucionaron a partir de los intrones.
2011-0119 dios te bendiga contuacion Maduración y Salida de Virus Desnudos y Envueltos
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. (Figura 1).
(B) Entada vía endosomas en la superficie celular (figura 2) Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Los antivirales son un tipo de medicamento usado para el tratamiento de infecciones producidas por virus. Tal como los antibióticos (específicos para bacteria), existen antivirales específicos para distintos tipos de virus. No sin excepciones, son relativamente inocuos para el huésped, por lo que su aplicación es relativamente segura. Deben distinguirse de los viricidas, que son compuestos químicos que destruyen las partículas virales presentes en el medio ambiente
Un virus satélite es un agente subvírico formado por ácido nucleico y que depende de la coinfección de la célula huésped por un virus (helper o máster) para reproducirse; obteniendo de ellos lasenzimas faltantes en su genoma, las cuales son necesarias para lograr replicarse.
Un virusoide o ARN satélite es una pequeña molécula patógena de ARN (ácido ribonucleico) que se encuentra encapsulada en ciertos virus de plantas con secuencias de ARN de mayor tamaño, lo que se denomina un virus asistente, el cual hospeda al virusoide y es necesario para que éste produzca una infección exitosa.
La McIntosh Roja o Mac es un tipo de manzana de piel roja y verde. Se recoge a finales de septiembre y es el cultivo más popular de Nueva Inglaterra, muy conocida por la compota rosada que se hace a partir de manzanas McIntosh sin pelar. Cada manzana de McIntosh tiene un linaje directo a un solo árbol descubierto en 1796 por John McIntosh en su granja en Dundela, una aldea situada en el condado de Dundas en la provincia canadiense de Ontario, cerca de Prescott.
Los plásmidos son fragmentos extracromosómicos de ácidos nucléicos (ADN o ARN) que aparecen en el citoplasma de algunos procariotas. Son de tamaño variable aunque menor que el cromosoma principal.
Los priones son los agentes causantes de un grupo de patologías neurodegenerativas letales características de mamíferos, también conocidas como encefalopatías espongiformes transmisibles. Estos agentes son capaces de propagarse dentro de un mismo huesped causando una lesión espongiótica y de transmitirse de huesped a huesped con elevados tiempos de incubación. A diferencia de virus y viroides, son resistentes a tratamientos inactivantes de ácidos nucléicos, pero comparten con éstos la existencia de una variabilidad de inóculos dentro de la misma especie (diferenciables por el patrón de la lesión y la magnitud del tiempo de incubación) y de una infectividad sujeta a barrera de especie (Chandler, 1961; Alper y cols., 1967; Hunter, 1972; Prusiner, 1982; Bruce y Fraser, 1991; Bessen y Marsh, 1992).
La búsqueda de la entidad molecular constitutiva de este agente reveló como componente mayoritario, si no único, una proteína: PrPSc, proteína del prion de scrapie),y la ausencia de un ácido nucleico específico (Prusiner 1982, 1991). Con estas premisas estructurales unidas a la capacidad de infección, Prusiner acuña el término prion (particula infecciosa de naturaleza proteica) para diferenciarlo de virus y viroides. Dadas las características poco convencionales de los priones se han elaborado numerosas hipótesis sobre su estructura (Dickinson y Outram, 1988; Prusiner, 1991; Weissmann, 1991). En la actualidad la hipótesis con mayor grado de aceptación es la conocida como "sólo proteína", delineada inicialmente por Griffith (1967) y, formalmente enunciada y actualizada por Prusiner (1991, 1997). Tras la descripción de proteínas de diferente secuencia pero similar comportamiento en levadura así como los avances interdisciplinares realizados en el conocimento de estos agentes en la última década, el concepto de prion ha sido acotado adquiriendo una defición más precisa y generalizable (Wicker y Masison, 1996; Lindquist S., 1997; Prusiner, 1997). Así, se denomina prion a la forma alterada de una proteína celular funcional (PrP en mamíferos) que ha podido perder su función normal pero que ha adquirido la capacidad de transformar la forma normal en patológica.
Han sido definidos como pequeñas partículas proteináceas que son resistentes a la inactivación por procedimientos que modifican los ácidos nucléicos.
Otra definición de prion es la forma alterada de una proteína celular funcional que ha podido perder su función normal pero que ha adquirido la capacidad de transformar la forma normal en patológica.
Los mamíferos tienen en su genoma genes que codifican la “proteína priónica”(PrP), y esta proteína en condiciones normales tiene una configuración espacial de tipo hélices alfa, que es susceptible a la lisis por proteasas y no es patógena. Los priones no son seres vivos ni tienen la capacidad de infección como tal.
La patogénesis de estas proteínas se produce cuando tienen bien una mutación (esporádica o adquirida genéticamente) o bien un cambio conformacional inducido por PrP-Sc (patógena) de otro individuo.
El cambio conformacional o las mutaciones hacen que la PrP tenga una estructura tridimensional diferente constituida por hélices beta con agregados amorfos insolubles (PrP-Sc), de manera que se hace imposible su lisis por las proteasas, produciéndose su acúmulo en los lisosomas de las células de los tejidos afectados y originando así las enfermedades priónicas.
Juan Miguel Mazara Simon 2011-0444 Martes, 3:00-5:00 PM
Virología es el estudio de virus y virus como agentes: clasificación y evolución, su estructura, sus formas de infectar y explotar las células para la reproducción del virus, las enfermedades que causan, las técnicas para aislar y la cultura de ellos y su uso en la investigación y terapia. Virología es considerada una parte de la microbiología o de patología. Una importante rama de la Virología es la clasificación de virus. Virus pueden clasificarse según la célula huésped que infectan: virus animales, virus de plantas, hongos virus y bacteriófagos (virus que infectan bacterias, que son los virus más complejos). Otra clasificación utiliza la forma geométrica de su cápside (a menudo una hélice o un icosaedro) o la estructura del virus (por ejemplo, la presencia o la ausencia de un envolvente de lípidos). Gama de virus en tamaño de unos 30 nm a aproximadamente 450 nm, lo que significa que la mayoría de ellos no puede verse con microscopios de luces. La forma y la estructura del virus ha sido estudiado por cristalografía de rayos X, microscopía electrónica y RMN. El sistema de clasificación más útil y más ampliamente utilizado distingue virus segun el tipo de ácido nucleico que utilizan como material genético y el método de replicación viral que emplean coaxial células huésped en la producción de más virus: Virus de ADN (divididas en virus de ADN bicatenario y los virus de ADN monocatenario mucho menos común), Virus ARN (dividieron en virus de ARN monocatenario de sentido positivo, virus de ARN monocatenario de sentido negativo y los virus de ARN bicatenario mucho menos común), invertir transcribir virus (virus de ADN de transcripción inversa de bicatenario y monocatenario transcripción inversa RNA virus incluyendo retrovirus). Además virólogos también estudian '' partículas subvirales '', incluso más pequeñas que los virus infecciosas entidades: viroides (desnudas circulares moléculas de ARN infectando plantas), satélites (moléculas de ácido nucleico con o sin una cápside que requieren un virus auxiliar para la infección y reproducción) y priones (proteínas que pueden existir en una conformación patológica que induce a otras moléculas del prión para asumir esa misma conformación). El último informe del Comité Internacional de taxonomía de virus (2005) muestra 5450 virus, organizados en más de 2.000 especies, 287 géneros y 73 familias 3 pedidos. Los taxones en Virología no son necesariamente monofiléticos. De hecho, las relaciones evolutivas de los distintos grupos de virus no están claras, y existen tres hipótesis sobre su origen: Virus surgieron de materia no viviente, por separado y en paralelo a otras formas de vida, posiblemente en forma de autorreplicación RNA ribozimas similares a viroides. Virus surgieron de la anteriores, más competentes celulares formas de vida que se convirtieron en parásitos a las células del huésped y posteriormente perdió la mayoría de sus funciones; ejemplos de tales diminutos parásitos procariotas son Mycoplasma y Nanoarchaeum.
Virus surgieron como partes del genoma de las células, probablemente transposones o plásmidos, que adquirieron la capacidad para "romper libre" desde el host de celda y infectan otras células. Por supuesto es posible que diferentes alternativas se aplican a grupos de virus diferente. De particular interés aquí es mimivirus, un virus gigante que infecta a las amebas y lleva gran parte de la maquinaria molecular tradicionalmente asociada con las bacterias. ¿Es una versión simplificada de una Célula procariota parasitaria o se originó como un simple virus que adquirido genes de su host? La evolución del virus, que a menudo se produce en concierto con la evolución de sus anfitriones, se estudia en el campo de la evolución viral. Mientras que el virus se reproducen y evolucionan, ellos no participan en el metabolismo y dependen de una célula huésped para la reproducción. La cuestión debatida a menudo de si están vivos o no es una cuestión de definición que no afectan la realidad biológica de los virus. Un virus (de la palabra latina virus, toxina o veneno) es una entidad biológica capaz de autorreplicarse utilizando la maquinaria celular, es un agente potencialmente patógeno compuesto por una cápside (o cápsida) de proteínas que envuelve al ácido nucléico, que puede ser ADN o ARN. Esta estructura puede a su vez estar rodeada por la envoltura vírica, una capa lipídica con diferentes proteínas, dependiendo del virus. El estudio de los virus ha sido muy complejo para los científicos, ya que ellos tienen la capacidad de cambiar permanentemente y esto dificulta su control y eliminación. Producen enfermedades como sarampión, gripe, viruela, poliomielitis y un mal que afecta a muchas personas en el mundo: el síndrome de inmuno deficiencia adquirida o sida. El ciclo vital de un virus siempre necesita de la maquinaria metabólica de una célula para poder replicar su material genético, produciendo muchas copias del virus original. En dicho proceso reside la capacidad destructora de los virus, ya que pueden perjudicar a la célula hasta destruirla. Pueden infectar células eucarióticas o procarióticas (en cuyo caso se les llama bacteriófagos, o simplemente fagos). Algunos indicios parecen demostrar que existen virus que infectan a otros virus (llamados viroides). Se puede agrupar las características definitorias de los virus en torno a tres cuestiones: su tamaño, el hecho de que sean cristalizables y el hecho de que sean parásitos intracelulares o microcelulares obligados. Estas tres cuestiones colocan a los virus en la frontera entre lo vivo y lo inerte. Tamaño de los virus Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus.
Su pequeño tamaño explica lo tardío del descubrimiento de estos agentes. La primera referencia sobre la existencia de los virus se debe al botánico ruso Dimitri Ivanovski en 1892. Este investigador buscaba el agente causante de la enfermedad denominada mosaico del tabaco, y llegó a la conclusión de que debía tratarse de una toxina o de un organismo más pequeño que las bacterias, pues el agente atravesaba los filtros que retenían las bacterias. Denominó a estos agentes patógenos virus filtrables. En 1897, el microbiólogo holandés Martinus Beijerink realizó experimentos similares a los de Ivanovski, y llegó a desechar la idea de las toxinas, pues se trataba de un agente capaz de reproducirse, ya que mantenía su poder infeccioso de unas plantas a otras, sin diluirse su poder patógeno. Poco después, los microbiólogos alemanes Frederick Loeffler y Paul Frosch descubrieron que la fiebre aftosa del ganado era producida por un virus filtrable que actuaba como un agente infeccioso. En la década de los 30, con el uso de filtros de tamaño de poro inferior, con las técnicas de cultivo celular in vitro que permitían la obtención de gran cantidad de virus, con la ultracentrifugación y finalmente con el microscopio electrónico y la difracción de rayos X, se logró visualizar a estos agentes. Cristalización Los virus son cristalizables, como demostró W. Stanley en 1935. Esto significa que las partículas víricas tienen formas geométricas y que son idénticas entre sí, lo cual las separa de la irregularidad característica de los organismos, y las acerca a las características de los minerales. Parásitos intracelulares obligados Los virus son parásitos intracelulares obligados. Desde los años treinta se sabe que los virus se componen principalmente de ácido nucleico y proteínas, estas últimas forman la cápside, que se conoce también como envoltura proteínica. Esto quiere decir que necesitan un huésped, ya que en vida libre no sobreviven. Se sabe que los virus pueden vivir alrededor de unos cuarenta días sin que tengan algún huésped en que se continúen reproduciendo. También se han encontrado virus que presentan lípidos, aunque estos son tomados de la célula que infectan. Hasta ahora todos los virus que se conocen presentan un solo tipo de ácido nucleico (ya sea ADN ó ARN), el cual puede ser de una ó de dos cadenas y puede ser segmentado. Para que el ácido nucleico pueda replicarse, necesita utilizar la maquinaria encimática y estructural de una célula viva, y por otra parte, solamente dentro de una célula viva tienen los virus las funciones de autoconservación, que junto con la reproducción, caracterizan a los seres vivos. Esta condición es la causa de que muchísimos virus sean conocidos como gérmenes patógenos que producen enfermedades en plantas y animales, e incluso en las bacterias. Estructura de los virus Un virus está compuesto de una molécula de ácido nucleico y una envoltura proteínica. Ésta es la estructura básica de un virus, aunque algunos de ellos pueden añadir a esto la presencia de alguna enzima, bien junto al ácido nucleico, como la transcriptasa inversa de los retrovirus, bien en la envoltura, para facilitar la apertura de una brecha en la membrana de la célula hospedadora.





















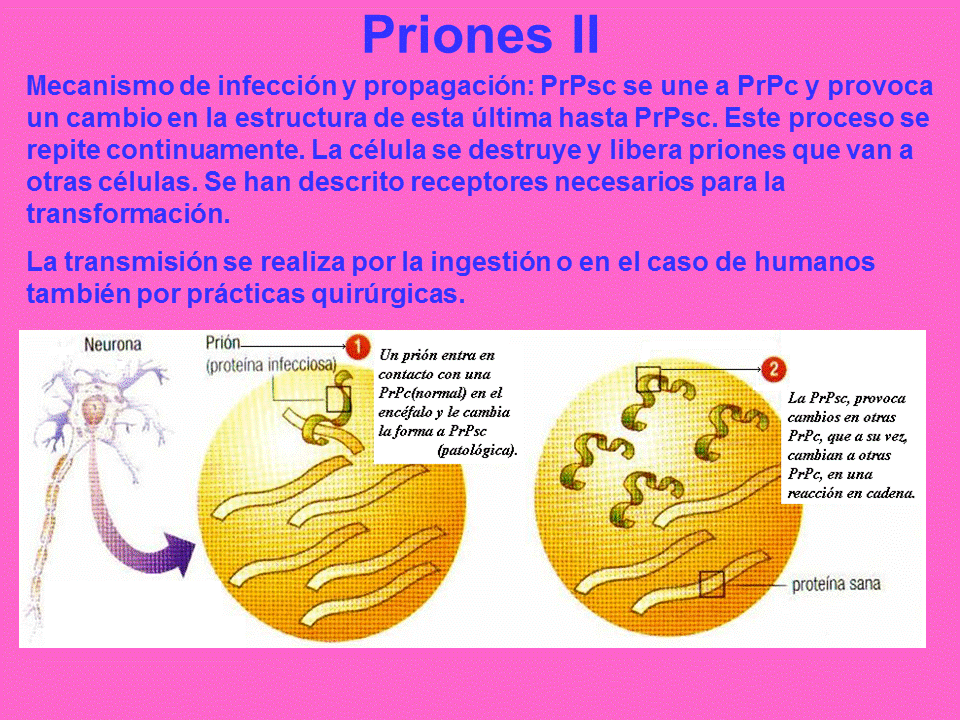
Origen de las células eucariotas (2011-0560)
ResponderEliminarEl origen de los eucariontes es un proceso que tiene un origen procariota. Este proceso se produjo por endosimbiosis entre varios organismos procariotas, en donde el ancestro principal protoeucariota es de tipo arqueano y las mitocondrias y cloroplastos son de origen bacteriano. Es discutible la incorporación de otros organismos procariotas. Las procariotas se diferenciaron de las eucariotas porque las procariotas no tienen núcleo y tienen genoma y las eucariotas si. Ejemplo de esto, es que las cianos bacterias entraron en procariotas grandes y la convirtieron en eucariotas, como las algas dieron origen a las plantas.
Este comentario ha sido eliminado por el autor.
ResponderEliminarEste comentario ha sido eliminado por el autor.
ResponderEliminarEste comentario ha sido eliminado por el autor.
ResponderEliminar2011-0757
ResponderEliminarColoración del Gram
La tinción de Gram o coloración de Gram es un tipo de tinción diferencial empleado en Bacteriología para la visualización de bacterias, sobre todo en es una prueba potente y rápida que nos permite diferenciar dos clases de bacterias.
Es una prueba potente y rápida que nos permite diferenciar dos clases de bacterias estas son: Bacterias gram positivas y las gram negativas.
Las grampositivas se tiñen de morado ya que el colorante se queda atrapad en la capa gruesa de peptidoglucanos que rodea a la célula
Las gramnegativas tienen una capa de petidoglucano mucho más delgada es por ello que no retiene el violeta cristal y por esto las células se tiñen con safranina y las observamos rojas.
2011-0757
ResponderEliminarFormas y agrupaciones bacteriana
Los principales tipos de formas bacterianas son:
*Cocos (células más o menos esféricas)
*Bacilos (en forma de bastón, alargados),que a su vez pueden tener varios aspectos:cilíndricos,fusiformes y en forma de maza, etc.
Atendiendo a los tipos de extremos, éstos pueden ser:
*Redondeados (lo más frecuente)
*Cuadrados
*Biselados
*Afilados
*Espirilos: al igual que los bacilos, tienen un eje más largo que otro, pero dicho eje no es recto, sino que sigue una forma de espiral, con una o más de una vuelta de hélice.
*Vibrios: proyectada su imagen sobre el plano tienen forma de coma, pero en el espacio suelen corresponder a una forma espiral con menos de una vuelta de hélice.
Otros tipos de formas son filamentos, ramificados o no,anillos casi cerrados, y formas con prolongaciones (con prostecas).
AGRUPACIONES BACTERIANAS
Las bacterias normalmente se multiplican por fisión transversal binaria. En muchas especies, las células hijas resultantes de un evento de división por fisión tienden a dispersarse por separado al medio, debido a la actuación de fuerzas físicas (movimiento browniano, cizallamiento, corrientes de convección, etc). Esto hace que al observar al microscopio una población de estas bacterias veamos mayoritariamente células aisladas. Pero en algunas especies las células hijas pueden permanecer unidas entre sí (al menos durante un cierto tiempo tras la división de la que proceden) debido a que el tabique sea incompleto o a la existencia de capas mucosas que retienen juntos los productos de la división.
Si la tendencia a permanecer unidas es baja, tendremos agrupaciones de dos células, que dependiendo que sean de morfología esférica o alargada, se denominan como:
*Diplococos
*Diplobacilos
Si la tendencia a permanecer unidas es mayor (por más tiempo), nos encontramos con varias posibilidades, dependiendo del número de planos de división y de la relación entre ellos:
Si los tabiques son paralelos entre sí (o sea, existe un solo plano de división): estreptococos (cadenetas arrosariadas de cocos) y estreptobacilos (cadenetas de bacilos).
Si existe más de un plano de división, en el caso de cocos podemos encontrar tres posibilidades:
Dos planos perpendiculares: tétradas (4 céls. en un plano) o múltiplos.
Tres planos ortogonales: sarcinas (paquetes cúbicos).
Muchos planos de división: estafilococos (racimos irregulares).
En el caso de bacilos, se pueden dar variantes adicionales, debido a la posibilidad de que se produzcan movimientos postfisionales (en algunos casos con desgarro):
Bacilos en empalizada o en paquetes de cigarrillos (debido a giros de 180grado)
Dos bacilos en ángulo (en forma de letra V o L)
Varios bacilos formando “letras chinas”.
2011-0757
ResponderEliminarAgrupación bacteriana
Algunos géneros bacterianos se agrupan de una manera característica. Esta agrupación se debe a la tendencia de las células hijas a permanecer parcialmente adheridas después de la división celular.
Los cocos pueden disponerse:
-De a pares y se los llama diplococos.
-Si se disponen en cadena se llaman estreptococos.
-Cuatro células esféricas conforman una tetrada
en forma de racimo o irrregular se llaman estafilococos.
-En paquetes cúbicos se denominan sarcinas.
MATRICULA:2011-0328 GRUPO:5
ResponderEliminar*LA VIROLOGIA(1)
Antes de hablar de lo que es la virologia debemos dar un pequeño concepto de lo que es un virus.
La palabra virus viene de veneno. Los agentes que hoy conocemos se designaron hace muchos años con el nombre de “agentes filtrables”. La visualización del virus se realizó en la cristalografía y comprobó la presencia de elementos lineales en series hexagonales que contienen ARN y proteínas. Las enfermedades producidas por virus han afectado a la humanidad desde años remotos.
La virología médica está presente en todas las especialidades y áreas médicas. Los virus son partículas formadas por ácido nucleico y proteínas. Considerados como la forma mas pequeña de vida, los virus, son partículas inertes que adquieren vitalidad y se reproducen cuando entran a una célula de un huésped susceptible, y no se multiplican por división, sino que una vez sintetizados sus componentes, estos se ensamblan.
La Virología es la materia que se ocupa del estudio de los virus.Etos son los causantes de numerosas enfermedades.
Los virus son importantes herramientas en investigación. Utilizando virus se ha avanzado en la Biología Molecular, conocimiento de genes, de mecanismos de replicación, trascripción, ác. nucleicos, etc. Los genomas de los virus son muy usados en Ingeniería Genética como vectores para transferir genes de unos individuos a otros.
Tienen importancia biológica. Todos los seres vivos del planeta tienen virus que los parasitan.
MATRICULA:2011-0328 GRUPO:5
ResponderEliminarCONTINUACION.
*ESTRUCTURA GENERAL DE LOS VIRUS.(2)
Como se puede observar en esta imagen todos los virus están constituidos por material genético DNA o RNA (pero no ambos) y una cápside formada por proteínas. Al conjunto del genoma y de la cápside se le denomina núcleo cápside, que puede estar rodeada o no de una envoltura. Podemos diferenciar varios tipos de virus en función de su constitución: desnudos o con envoltura.
*Los virus desnudos, según el tipo de cápside, se diferencian en 3 tipos:
-Icosaédricos. La cápside es un icosaedro.
-Helicoidales. La cápside es un cilindro hueco.
-Complejos. Tienen diferentes estructuras anejas a la cápside (cola,..).
*Los virus con envoltura se diferenciar también en tres tipos:
-Icosaédricos.
-Helicoidales.
-Complejos.
Podemos definir su composición de la siguiente manera:
-Ácidos nucleicos virales:
Los virus se caracterizan, a diferencia de los otros organismos, por presentar una única especie de ácido nucleico constitutiva que puede ser ADN o ARN, monocatenario o bicatenario con estructura de doble hélice.
-Cápsides:
La cápside es una cubierta proteica externa que encierra y protege al genoma viral de la acción de nucleasas y otros factores adversos del medio exterior. Además, en los virus desnudos carentes de envoltura, la cápside es la encargada de establecer a través de alguna de sus proteínas la unión con la célula que será parasitada por el virus. Asimismo, las proteínas de la cápside contienen los determinantes antigénicos contra los que el sistema inmune del huésped elaborará la respuesta de anticuerpos en defensa del organismo.
-Envolturas
La envoltura de un virus es una membrana constituida por una doble capa lipídica asociada a glicoproteínas que pueden proyectarse en forma de espículas desde la superficie de la partícula viral hacia el exterior.
Los virus adquieren su estructura mediante un proceso de brotación a través de alguna membrana celular. El número de glicoproteínas que presentan los virus animales es muy variable.
MATRICULA:2011-0328 GRUPO:5
ResponderEliminarCONTINUACION
*SIMETRIA VIRAL.(3)
La simetría es la forma que adopta un virus en el espacio y está dad por la estructura de su nucleocápside.
Como se puede observar en la siguiente imagen la simetría puede ser helicoidal, icosaédrica ,binaria o compleja.
-Los virus con simetría icosaédrica son poliedros regulares con 20 caras triangulares, 30 aristas y 12 vértices. Son ejemplos de virus con simetría icosaédrica desnudos el virus de la poliomielitis o los adenovirus. Los herpesvirus y togavirus son virus con simetría icosaédrica y recubiertos por una envoltura.
-Los virus con simetría helicoidal presentan una nucleocápside cilindrica que puede estar extendida como en el virus del mosaico del tabaco. Este es un virus desnudo. En el caso de la imagen que tenemos aquí la nucleocápside está arrollada sobre sí misma y recubierta por una envoltura, como en el virus Influenza.
-los virus con simetría binaria tienen cabeza icosaédrica y cola hélica. El ácido nucleico se encuentra en la cabeza. Bacteriófagos.
-Se denominan virus de simetría compleja a aquellos que presentan una nucleocápside helicoidal o icosaédrica recubierta por una envoltura laxa. Ejemplos de éstos virus son los poxvirus (forma de ladrillo), los rhabdovirus (forma de bala) y algunos bacteriófagos que poseen una cabeza con simetría icosaédrica y una cola con simetría helicoidal.
*TAMAÑO DE LOS VIRUS.(4)
En el siguiente recuadro notamos la variedad de tamaños con respecto a las células que están en comparación.
Se puede decir que los virus son estructuras extraordinariamente pequeñas.
La mayoría de los virus poseen un tamaño muy inferior al de las bacterias. Por ello, sólo pueden observarse con el microscopio electrónico.
Dado su pequeño tamaño es necesario utilizar para su medición una medida inferior al micrón, denominada nanómetro (nm) o milimicrón (mu) que equivales a 10-9 metros.
Los virus mas pequeños, por ejemplo los que integran la familia Picornaviridae, donde se incluye el virus de la poliomielitis, miden alrededor de 27 nm y los más grandes, como los de la familia Poxviridae, donde se encuentra el virus de la viruela, miden aproximadamente 250 nm.
La mayoría de los virus estudiados tienen un diámetro de entre 10 y 300 nanómetros. Algunos Filovirus tienen un tamaño total de hasta 1.400 nm, sin embargo, sólo miden unos 80 nm de diámetro.La mayoría de virus no pueden ser observados con un microscopio óptico, de manera que se utilizan microscopios electrónicos de barrido y de transmisión para visualizar partículas víricas.
MATRICULA:2011-0328 GRUPO:5
ResponderEliminarCONTINUACION
FAMILIAS DE VIRUS.(5)
(patogenos para el hombre)
Aquí se puede notar una gran diversidad de estos entre los cuales podemos mencionar:
-PicoRNAvirus:
Por picornavirus se conoce a todo virus que pertenece a la familia Picornaviridae. Los picornavirus son virus no envueltos, de única cadena ARN con cápside icosaédrica.
Su genoma es inusual ya que tiene una proteína en el extremo 5 ‘que se utiliza como cebador para la transcripción por la ARN polimerasa. El nombre se deriva de “pico”, que significa pequeño, ya que es literalmente un pequeño virus de ARN.
Los picornavirus se separan en una serie de géneros e incluyen muchos importantes patógenos de humanos.
Las enfermedades que causan son muy variadas, que van desde aguda “gripe común” a enfermedades como la poliomielitis, y otras infecciones crónicas.
- Los Calicivirus:
Son Virus RNA pequeños pertenecientes a la familia Caliciviridae. Dentro de esta familia destacan dos géneros: los Norovirus, también conocidos como "virus tipo Norwalk" o "virus pequeños redondos estructurados" y los Sapporovirus.
- Hepadnavirus:
Los hepadnavirus son un subgrupo de los virus que producen hepatitis.
Es uno de los virus más importantes desde el punto de vista médico. Infecta a más de un cuarto de billón de personas y es una de las principales causas de cirrosis y cáncer hepático (carcinoma hepatocelular). HBV, también representa el primer ejemplo del éxito de una vacuna recombinante para una enfermedad contagiosa humana, y puede tener importantes implicaciones para otras enfermedades de transmisión sexual, como el HIV.
- Reovirus:
Grupo de tres virus de doble cadena de ARN presentes en todas partes, identificados en los aparatos respiratorio y digestivo tanto de personas sanas como de enfermas. Los reovirus han sido implicados en algunos casos de enfermedades de vías respiratorias superiores y de gastroenteritis infantiles.
- Los Rhabdovirus:
Son virus con cápside helicoidal. Incluye al virus de la rabia y de la estomatitis vesicular (que es el organismo modelo). El virus de la estomatitis vesicular es un virus de 180x75nm, con cápsida rodeada por una envoltura con proyecciones constituídas por la proteína G.
- Togavirus:
Familia de arbovirus a la que pertenecen los organismos causantes de encefalitis, del dengue, de la fiebre amarilla y de la rubéola.
- Coronavirus:
Género de virus de la familia de los Coronaviridae El coronavirus humano es el responsable de infecciones respiratorias de las vías superiores. Miembro de la familia de los virus que abarcan distintos tipos capaces de producir una enfermedad respiratoria aguda.
- Bunyavirus:
Género de virus ARN de vertebrados de la familia Bunyaviridae.
-Papovavirus:
Grupo de pequeños virus ADN, algunos de los cuales pueden ser potencialmente productores de cáncer. La verruga humana está producida por un tipo de papovavirus. Algunos tipos de papovavirus son: papovavirus del papiloma, papovavirus del polioma y papovavirus SV.
- Paramixovirus:
Miembro de una familia de virus que abarca a los organismos causantes de la gripe, de las paperas y de algunas infecciones respiratorias.
- Retrovirus:
Virus de ácido ribonucleico que posee una enzima que le permite convertir su material genético en ácido desoxirribonucleico cuando ha penetrado en una célula viva. Al grupo de los retrovirus pertenece el virus del sida.
- Filovirus:
Género de virus con ácido ribonucleico de cadena simple con sentido negativo, de la familia Filoviridae. El género incluye los virus Ébola y Marburg.
- Poxvirus:
Miembro de una familia de virus que incluye a los organismos que producen el molusco contagioso, la viruela y la vacuna.
MATRICULA:2011-0328 GRUPO:5
ResponderEliminarCONTINUACION
*DIAGNOSTICO DE LAS VIROSIS.(6)
Antes de hablar de diagnostico es bueno hacernos esta pregunta:
¿Cómo infectan los virus al ser humano?
Los virus pueden penetrar en el cuerpo humano por cualquiera de sus “puertas de entrada,” pero por lo general entran por la nariz y la boca. Una vez dentro del cuerpo, el virus se adhiere a la superficie de la célula que piensa atacar, denominada célula huésped.
Una vez dentro, comienza a elaborar otros virus idénticos a partir de la proteína de la célula huésped. Los nuevos virus pueden salir a través de la membrana externa de la célula huésped, a la que a veces destruyen antes de abandonarla, y se lanzan al ataque contra otras células huésped. El proceso prosigue hasta que el cuerpo elabora suficiente cantidad de anticuerpos y otras defensas para derrotar a los virus invasores.
-Los síntomas son muy variables y dependen del virus y del órgano afectado. Muchos virus, como muchas bacterias, producen fiebre y síntomas respiratorios (tos, estornudos) o intestinales (náuseas, vómito, diarrea). Las virosis, aun cuando no sean de peligro, a menudo provocan fiebre elevada en los niños de corta edad.
- Algunas infecciones virales, como la gripe, el resfriado comúno, la varicela son fáciles de identificar por sus síntomas, sin que se necesiten análisis de laboratorio. Para otras, como la hepatitis viral, el sida y la mononucleosis infecciosa, se suele analizar una muestra de sangre en busca de anticuerpos específicos contra el virus. La presencia de estos anticuerpos confirma el diagnóstico.
- En ciertos casos, los virus pueden cultivarse en tejidos celulares o bien el virus se identifica por su ácido nucleico, valiéndose de una técnica que recibe el nombre de reacción en cadena de la polimerasa (RCP). Las pruebas como el RCP o el cultivo celular se emplean cuando las de anticuerpos no tienen la precisión suficiente o cuando es necesario cuantificar con mayor exactitud el virus invasor.
*ETAPAS DE LA REPLICACION VIRAL.(7)
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
MATRICULA:2011-0328 GRUPO:5
ResponderEliminarCONTINUACION
*ADHESION.(8)
El primer paso en la infección de una célula es la adhesión o fijación a la superficie celular. Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
*PENETRACION.(9)
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
-Virus envueltos.( Figura B)
1. Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
2.Entrada vía endosomas en la superficie celular.
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
-Virus no envueltos o desnudos (Figura A)
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
*PENETRACION DE LOS BACTERIOFAGOS.(10)
Los bacteriófagos tienen la modalidad de adsorber y penetrar, mejor conocida por su mecanismo de inyección. Las proteínas que forman las espinas de la placa basal entran en contacto mediante sus receptores con los de la pared bacteriana y se anclan a ella.
La accion de algunas enzimas hidroliticas de la capside rompe la pared y esta es perforada por el eje tubular.
El genoma se introduce por el eje hasta el citoplasma de la bacteria quedando fuera de la celula la capsida. Los virus vegetales pasan la pared celular a través de pequeñas grietas producidas por picaduras de insectos.
Los virus animales tienen acceso mas directo a la membrana de las celulas. Los virus sin envoltura penetran por completo en la célula por endocitosis y una vez dentro de la vesicula,el virus libera enzimas hidroliticas para romper la vesicula, y quedar libre en el citoplasma el virus.
Los virus con envoltura penetran al fusionar su cubierta con la membrana celular,y se forma una vesicula endocítica que se une a un endosoma,cuya membrana se fusiona con la envoltura del virus,escapando de ser digerido en su interior.
*REPLICACION Y MADURACION.(11)
La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínas víricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores. Los ribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada.
Como podemos observar en la imagen:
-Los virus con ADN realizan la replicación del material genético de la misma manera que las células; en el caso de los virus con ADN monocatenario, previamente a la replicación se sintetiza una cadena de ADN complementario para formar la doble hélice.
-Los virus con ARN replican el material genético sin necesidad de pasar por ADN, actuando cada cadena de ARN como molde para la síntesis de su complementaria.
MATRICULA:2011-0328 GRUPO:5
ResponderEliminarCONTINUACION
*MADURACION Y SALIDA DE VIRUS DESNUDOS Y ENVUELTOS.(12)
.
El desnudamiento ocurre simultáneamente o después de la penetración y comprende la eliminación de todas o parte de las cubiertas proteicas que rodean al ácido nucleico viral, de manera de dejar expuesto dentro de la célula el genoma viral desnudo o asociado a alguna enzima para que pueda expresarse.
En algunos casos sólo se remueve parte de la cápside viral y en esas condiciones el genoma expresa sus funciones. Para otros virus complejos, como por ejemplo los Poxvirus, el desnudamiento parece ocurrir en dos etapas: en la primera hay remoción de ciertas cubiertas proteicas por enzimas celulares, parte del genoma es tanscripto en esas condiciones y luego, en una segunda etapa, y a través de una proteína codificada por el virus, se produce el desnudamiento total del genoma.
En general el desnudamiento es un proceso difícil de estudiar y establecer con precisión sus características por la rapidez con que ocurre y la dificultad en separarlo temporalmente de la penetración. Es por ello que se suele denominar internalización del virión al conjunto de ambos eventos, penetración y desnudamiento.
*SALIDA DE UN VIRUS ENVUELTO.(13)
Vemos aquí como los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes.
No todas las partículas víricas liberadas son infecciosas.
La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
Los virus envueltos (como el VIH) son liberados de la célula huésped por gemación. Durante este proceso, el virus adquiere su envoltura, que es una parte modificada de la membrana plasmática del huésped.
MATRICULA:2011-0328 GRUPO:5
ResponderEliminarCONTINUACION
*ANTIVIRALES.(14)
Los farmacos que inhiben procesos especificos del virus, como la union a la celula, la decapsidacion del genoma viral o el ensamblaje de los nuevos virus, o, preferentemente, inhiben la sintesis macromolecular dirigida por el virus.
Actualmente se dispone de farmacos antivirales de probada eficacia terapeutica y profilactica para varias infecciones virales comunes y algunas potencialmente mortales. Ademas, continua la busqueda de nuevos farmacos antivirales y nuevas estrategias terapeuticas para el tratamiento de las enfermedades virales.
En el siguiente recuadro se presenta una clasificacion de estos antivirales de la siguiente manera:
-Antivirales que impiden la entrada del virus a la celula:
•Amantadina: actua sobre el virus influenza A. Su efecto principal es inhibir la decapsidacion viral o el desensamblaje del virion durante la endocitosis.
•Rimantadina: derivado 4-10 veces mas activa que la amantadina.
•Oseltamivir: inhibe la actividad de neuraminidasa del virus influenza.
•Zanamivir:inhibe la actividad de neuraminidasa del virus influenza.
•Docosanol: interfiere en la fusion del VHS-1 y VHS-2 a las membranas de las celulas del huesped. Es de uso topico.
-Antivirales que actuan sobre la fase de replicación del genoma viral.
•Aciclovir: inhibe la sintesis del ADN viral. Tiene accion sobre los herpesvirus.
•Valaciclovir: profarmaco del Aciclovir.
•Ganciclovir: inhibe la sintesis del ADN viral. Tiene accion sobre los herpesvirus, pero su caracteristica singular es una potente inhibicion de la replicacion del CMV.
•Valganciclovir: profarmaco del Ganciclovir.
•Adefovir: Actua como inhibidor competitivo de las polimerasas del ADN y transcriptasas inversas virales con respecto al dATP. Se selectividad se relaciona con una mayor afinidad por la ADN polimerasa del VHB que por las polimerasas de las celulas huesped. Esta aprobado para el tratamiento de la hepatitis B cronica.
•Cidofovir: inhibe la sintesis de ADN viral de los herpesvirus.
MATRICULA:2011-0328 GRUPO:5
ResponderEliminarCONTINUACION
*MECANISMO DE ACCION DE LOS RETROVIRALES.(15)
Con esta imagen podemos describirlo d ela siguiente manera y tomare como ejemplo el virus del VIH:
-Inhibidores de la entrada
La infección por el VIH se inicia con la unión de la partícula viral a la membrana externa de las células diana, mediante la interacción entre la gp120 del VIH y dos moléculas celulares: el receptor CD4 y el correceptor de quimocinas (CCR5 o CXCR4). En un paso posterior, la gp41 viral se adhiere a la superficie celular para iniciarse la fusión de las membranas del VIH y de la célula diana. Para que esto ocurra es preciso que la gp41 se pliegue sobre sí misma acortando el espacio que separa ambas membranas.
-Inhibidores de transcriptasa inversa
Se trata de fármacos que inhiben la síntesis del ADN complementario a partir del ARN viral, función realizada por la transcriptasa inversa (TI) viral en el interior de la célula infectada. Según el mecanismo de bloqueo de la TI se dividen en:
-Inhibidores de la integrasa
Una vez sintetizado el ADN complementario, éste pasa al interior del núcleo de la célula para allí integrarse en el ADN celular. La integrasa viral es la enzima responsable de este proceso. Actualmente se encuentran en desarrollo clínico una serie de compuestos que bloquean el ciclo viral al nivel de la integración del ADN complementario en el ADN celular.
-Inhibidores de la proteasa
Como se ha señalado, el ADN complementario viral sintetizado por acción de la TI se integra en el ADN de la célula infectada. El ciclo viral continúa cuando el ADN viral se transcribe a ARN mensajero que en los ribosomas celulares se traduce en una poliproteína viral. Esta cadena proteica ha de escindirse en diversos péptidos virales que dan lugar a las proteínas estructurales y funcionales que formarán parte de las nuevas partículas virales. La proteasa viral es la enzima responsable de la hidrólisis de la poliproteína en los diversos péptidos individuales. Existe un grupo de fármacos, los inhibidores de la proteasa (IP), que basan su mecanismo de acción en el bloqueo de esta enzima. De este modo se impide la maduración y ensamblaje de las partículas virales, y la infección de nuevas células diana.
*RETROVIRALES.(16)
Entre las clases de retrovirus que observamos aquí podemos mencionar los siguientes:
-Inhibidores nucleosídicos de la transcriptasa inversa. Actúan sobre la enzima transcriptasa inversa, incorporándola a la cadena de ADN que crea el virus. Con ello, esa cadena se vuelve defectuosa e impiden que el virus se reproduzca.
Son los siguientes: Zidovudina, abacavir, didanosina, estavudina, lamivudina y tenofovir.
-Inhibidores no nucleósidos de la transcriptasa inversa. Bloquean directamente la acción de la enzima y la multiplicación del virus.
Son los siguientes: efavirenz, nevirapina y etravirina.
-Inhibidores de la proteasa. Actúan sobre la enzima proteasa, bloqueando su acción e impidiendo la producción de nuevas copias de células infectadas por el VIH.
Son los siguientes: amprenavir, atazanavir, darunavir, indinavir, lopinavir/r, nelfinavir, ritonavir y saquinavir.
-Inhibidores de la fusión. Impiden la entrada de virus a la célula, con lo que no puede reproducirse.
La enfuvirtida pertenece a este grupo.
-Inhibidores de la integrasa. Bloquean la actividad de la enzima integrasa, responsable de la inserción del ADN del VIH en el ADN humano (código genético de la célula). De esta manera inhibe la replicación del virus y su capacidad de infectar a nuevas células.
MATRICULA:2011-0328 GRUPO:5
ResponderEliminarCONTINUACION
*OTROS BIONES ACELULARES.(20)
-Satelites(schneider): Un satélite es un agente subvírico formado por ácido nucleico y que depende de la coinfección de la célula huésped por un virus (helper o máster). Si el satélite codifica una proteína de cubierta en la que se introduce el ácido nucleico, hablamos de virus satélite. Aunque su genoma no guarda relación con el del helper, sí que depende de las enzimas de éste para replicarse.
Un ejemplo es el virus satélite del mosaico del tabaco, que utiliza al virus del mosaico del tabaco para llevar a cabo su reproducción
- virusoides o sateloides(fransky): aunque son estudiados en virología, no se les considera verdaderos virus sino partículas subvirales, por necesitar de otros virus para propagarse. Por esto, en las clasificaciones de virus aparecen con el término de "ARN satélite circular". De hecho, el término virusoide ha sido también usado genéricamente para referirse a todos los ácidos nucleicos satélites.
- Los plásmidos : son moléculas de ADN extracromosómico circular o lineal que se replican y transcriben independientes del ADN cromosómico. Están presentes normalmente en bacterias, y en algunas ocasiones en organismos eucariotas como las levaduras. Su tamaño varía desde 1 a 250 kb. El número de plásmidos puede variar, dependiendo de su tipo, desde una sola copia hasta algunos cientos por célula.
MATRICULA:2011-0328 GRUPO:5
ResponderEliminarCONTINUACION
*PRIONES I .(21)
Los priones son los agentes causantes de un grupo de patologías neurodegenerativas letales características de mamíferos, también conocidas como encefalopatías espongiformes transmisibles. Estos agentes son capaces de propagarse dentro de un mismo huesped causando una lesión espongiótica y de transmitirse de huesped a huesped con elevados tiempos de incubación.
Los priones se propagan mediante la transmisión de proteínas anómalas con mal plegamiento. Cuando un prion entra en un organismo sano, actúa sobre la forma normal del mismo tipo de proteína existente en el organismo, modificándola y convirtiéndola en prion. Estos priones recién formados pueden pasar a convertir más proteínas, provocando una reacción en cadena que produce grandes cantidades de la proteína prion. Todos los priones conocidos inducen la formación de amiloides plegado, en los que actúan polimerasas formando un agregado que consiste en apretadas hojas ß.
El período de incubación de las enfermedades priónicas se determina por la tasa de crecimiento exponencial asociados con la replicación de priones, que es un equilibrio entre el crecimiento lineal y la rotura de los agregados (hay que tener en cuenta que la propagación del prion depende de la presencia de la proteína normalmente plegada en la que los priones pueden inducir plegamiento. Los organismos que no expresan la forma normal de la proteína priónica no pueden desarrollar o transmitir la enfermedad.
*PRIONES II. (22)
No se conocen con exactitud los mecanismos a través de los cuales la Prpc pasa a ser PrpSc. Una de las hipótesis es la rotura de los puentes de disulfuro entre aminoácidos de la estructura de hélice alfa (normalmente entre cisteínas que es un tipo de aminoácido que puede formar dichos puentes), hecho que da lugar a radicales –SH, presentes en las láminas beta.
Según las hipótesis actuales, los aminoácidos que actúan en el cambio conformacional son la metionina, por su capacidad de cambiar el radical metilo por –H consiguiendo un radical -SH; la cisteína, por su disposición a formar puentes disulfuro; y la valina, por su proximidad atómica a la Met y Cys.
En el laboratorio podemos romper los puentes disulfuro mediante dos mecanismos: a partir de la oxidación con ácido perfórmico o mediante la rotura con beta-mercaptoetanol y posterior acetilación con iodoacetato.
Otras hipótesis señalan que la sustitución del aminoácido prolina por leucina puede ser el posible responsable de la desestabilización de la estructura terciaria de la proteína y su transformación en agente patógeno, hecho apoyado con la comparación del gen PRNP procendente de un paciente enfermo con el de un paciente sano.
MATRICULA: 2011-0546 GRUPO:5
ResponderEliminarVirologia diap #1
Virología rama de la microbiología y Estudio de los virus y las enfermedades que éstos causan.
Estructura de los virus diap#2
Los virus más pequeños y simples están constituidos únicamente por ácido nucleico y proteínas. El ácido nucleico es el genoma viral, ubicado en el interior de la partícula, y puede ser ADN o ARN.Este conjunto de genoma y proteínas íntimamente asociadas es llamado "core", núcleo, nucleoproteína o nucleoide. Este núcleo central está rodeado por una cubierta proteica, la cápside, que junto con el genoma constituye la nucleocápside.
La cápside es una cubierta proteica externa que encierra y protege al genoma viral de la acción de nucleasas y otros factores adversos del medio exterior.
La envoltura de un virus es una membrana constituida por una doble capa lipídica asociada a glicoproteínas que pueden proyectarse en forma de espículas desde la superficie de la partícula viral hacia el exterior.
Los virus se caracterizan, a diferencia de los otros organismos, por presentar una única especie de ácido nucleico constitutiva que puede ser ADN o ARN, monocatenario o bicatenario con estructura de doble hélice.
MATRICULA: 2011-0546 GRUPO:5
ResponderEliminarSimetria de los virus diap#3
La simetría es la forma que adopta un virus en el espacio y está dad por la estructura de su nucleocápside. La simetría puede ser helicoidal, icosaédrica o compleja.
Los virus con simetría helicoidal presentan una nucleocápside cilindrica que puede estar extendida como en el virus del mosaico del tabaco. Este es un virus desnudo. En otros casos la nucleocápside está arrollada sobre sí misma y recubierta por una envoltura, como en el virus Influenza.
Los virus con simetría icosaédrica son poliedros regulares con 20 caras triangulares, 30 aristas y 12 vértices. Son ejemplos de virus con simetría icosaédrica desnudos el virus de la poliomielitis o los adenovirus. Los herpesvirus y togavirus son virus con simetría icosaédrica y recubiertos por una envoltura.
Se denominan virus de simetría compleja a aquellos que presentan una nucleocápside helicoidal o icosaédrica recubierta por una envoltura laxa. Ejemplos de éstos virus son los poxvirus (forma de ladrillo), los rhabdovirus (forma de bala) y algunos bacteriófagos que poseen una cabeza con simetría icosaédrica y una cola con simetría helicoidal.
Tamaño de los virus diap #4
Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus.
pero en realidad los tamaños de los virus se presentan de forma variable.
Familia de virus diap#5
Familia Especies importantes2
Adenoviridae adenovirus
Picornaviridae virus de hepatitis A, poliovirus
Herpesviridae Virus varicela-zoster
Hepadnaviridae virus de hepatitis B
Flaviviridae virus de hepatitis C
Retroviridae Virus de inmunodeficiencia humana (VIH)
Orthomyxoviridae virus de influenza
Paramyxoviridae virus virus del sarampión,
Papovaviridae
Rhabdoviridae virus de la rabia
Togaviridae virus de Rubéola
MATRICULA: 2011-0546 GRUPO:5
ResponderEliminarDiagnostico de la virosis diap #6
Podemos diagnosticar la virosis mediante metodos. Los métodos utilizados para reconocer las infecciones por virus humanos pueden clasificarse en DIRECTOS e INDIRECTOS
METODOS DIRECTOS
Son aquellos que detectan:
1. El virus como agente infeccioso (aislamiento viral):La base del diagnóstico viral es la detección del virus o de sus componentes. El aislamiento del virus era la técnica standard de oro sobre la cual se medían todas las otras pruebas de diagnóstico viral.
2. La presencia de antígenos virales (técnicas inmunológicas): Inmunofluorescencia (IF), Enzimoinmunoanálisis (EIA), Test de Aglutinación: (!)Esta técnica se puede utilizar para una identificación rápida del virus directamente sobre la muestra. (2) El test de aglutinación ha sido usado para detectar antígeno de Rotavirus en heces
3.La presencia de ácidos nucleicos virales (PCR, etc.).
4.El virus como partícula viral (microscopía electrónica).
MATRICULA: 2011-0546 GRUPO:5
ResponderEliminarEtapas de la replicacion viral diap#7,8,9,10,11,12,13
Dentro de estos podemos mencionar:
1). ADHESION: Esta adhesión se da vía interacciones iónicas las cuales son independientes de temperatura. La proteína de adhesión viral reconoce receptores específicos, los cuales pueden ser proteínas, carbohidratos o lípidos, en el exterior de la célula. Las células que carecen de los receptores apropiados no son susceptibles al virus.
2).PENETRACIÓN Y PENETRACION DE LOS BACTERIOFAGOS
Los virus penetran las células de maneras diversas dependiendo de la naturaleza misma del virus.
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular.
(B) Entada vía endosomas en la superficie celular
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
3). REPLICACION Y MADURACION (MADURACION Y SALIDA DE VIRUS Y ENVUELTOS)
El ácido nucleido debe de estar lo suficientemente expuesto como para que la replicación viral comience en esta fase. Cuando el ácido nucleico es expuesto, las partículas víricas infecciosas no pueden removerse de las células – este es el comienz de la fase de ECLIPSE – la cual perdura hasta que viriones infecciosos nuevos sean creados.Nuevas partículas víricas son ensambladas. Puede haber un estadio de maduración posterior al proceso inicial de ensamblaje.
4). SALIDA DE UN VIRUS ENVUELTO
Los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes. No todas las partículas víricas liberadas son infecciosas. La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
Antivirales diap#14
ResponderEliminarSe pueden clasificar en tres grandes grupos a los agentes quimioterapicos activos frente a los virus:
grupo 1:
Farmacos Viricidas: farmacos que directamente inactivan los virus intactos: Detergentes, Disolventes organicos: eter, cloroformo, Luz ultravioleta, Podofilinas, las cuales destruyen ademas tejidos del huesped.
Farmacos Antivirales: farmacos que inhiben procesos especificos del virus, como la union a la celula, la decapsidacion del genoma viral o el ensamblaje de los nuevos virus, o, preferentemente, inhiben la sintesis macromolecular dirigida por el virus.
Farmacos Inmunomodulares: comprenden los que reemplazan las respuestas inmunitarias deficientes del huesped y los que favorecen a los endogenos.
grupo 2: ANTIVIRALES QUE IMPIDEN LA ENTRADA DEL VIRUS A LA CELULA
Amantadina: actua sobre el virus influenza A. Su efecto principal es inhibir la decapsidacion viral o el desensamblaje del virion durante la endocitosis.
Rimantadina: derivado 4-10 veces mas activa que la amantadina.
Oseltamivir: inhibe la actividad de neuraminidasa del virus influenza.
Zanamivir:inhibe la actividad de neuraminidasa del virus influenza.
Docosanol: interfiere en la fusion del VHS-1 y VHS-2 a las membranas de las celulas del huesped.
grupo 3:ANTIVIRALES QUE ACTUAN SOBRE LA FASE DE REPLICACIÓN DEL GENOMA VIRAL
Aciclovir: inhibe la sintesis del ADN viral. Tiene accion sobre los herpesvirus.
Valaciclovir: profarmaco del Aciclovir.
Ganciclovir: inhibe la sintesis del ADN viral. Tiene accion sobre los herpesvirus, pero su caracteristica singular es una potente inhibicion de la replicacion del CMV.
Valganciclovir: profarmaco del Ganciclovir.
Adefovir: Actua como inhibidor competitivo de las polimerasas del ADN y transcriptasas inversas virales con respecto al dATP. Se selectividad se relaciona con una mayor afinidad por la ADN polimerasa del VHB que por las polimerasas de las celulas huesped. Esta aprobado para el tratamiento de la hepatitis B cronica.
Cidofovir: inhibe la sintesis de ADN viral de los herpesvirus.
Penciclovir: inhibe la sintesis de ADN viral de los herpesvirus.
Famciclovir. Profarmaco del Penciclovir de administracion oral.
Fomivirseno. Inhibe la replicacion del CMV humano. Se administra de forma intravitrea.
Foscarnet: actua sobre los herpesvirus y la transcriptasa reversa del HIV. Es inhibidor frente a la mayoria de las cepas de CMV resistentes a ganciclovir y de VHS y VVZ resistentes a aciclovir.
Idoxuridina. Inhibe la sintesis de ADN viral. Actua en especial sobre los herpesvirus y poxvirus.
Ribavirina: se utiliza en el tratamiento de la infección por HCV y por el VSR.
Trifluridina.
Vidarabina.
MATRICULA: 2011-0546 GRUPO:5
ResponderEliminarMecanismo de accion de los retrovirales diap#15 y 16
Los retrovirus son pequeños microorganismos que contienen una única cadena de ARN. Su ciclo vital se basa en la conversión de su ARN geonómico en doble cadena de ADN y posteriormente en la integración de éste último en los cromosomas de la célula huésped en lo que se denomina provirus endógeno.
Entre las clases de retrovirus que observamos aquí podemos mencionar los siguientes:
-Inhibidores nucleosídicos de la transcriptasa inversa. Actúan sobre la enzima transcriptasa inversa, incorporándola a la cadena de ADN que crea el virus. Con ello, esa cadena se vuelve defectuosa e impiden que el virus se reproduzca.
-Inhibidores no nucleósidos de la transcriptasa inversa. Bloquean directamente la acción de la enzima y la multiplicación del virus.
-Inhibidores de la proteasa. Actúan sobre la enzima proteasa, bloqueando su acción e impidiendo la producción de nuevas copias de células infectadas por el VIH.
-Inhibidores de la fusión. Impiden la entrada de virus a la célula, con lo que no puede reproducirse.
-Inhibidores de la integrasa. Bloquean la actividad de la enzima integrasa, responsable de la inserción del ADN del VIH en el ADN humano (código genético de la célula). De esta manera inhibe la replicación del virus y su capacidad de infectar a nuevas células.
MATRICULA: 2011-0546 GRUPO:5
Biones Acelulares diap#17
Los biones celulares son aquellos que se replican. Entre estos encontramos:
Viroides 1 y 2 (#18 y 19):
- Pequeñas moléculas de ARN circular de cadena sencilla formadas por unos pocos cientos de nucleótidos. Carecen de
cápsida y su tamaño es una milésima parte del de los virus más pequeños.
- Aparecen con mayor abundancia en el núcleo de las células infectadas.
- Sólo han sido detectados en plantas, en las que producen una gran variedad de enfermedades y pueden ser transmitidos tanto horizontal como verticalmente.
- No existen ninguna evidencia de que los viroides son traducidos a proteínas, ni se conoce cómo causan las enfermedades. Lo que sí se sabe es que son replicados por los enzimas del huésped.
- Algunas semejanzas entre la secuencia de nucleótidos de los viroides y ciertos secuencias del genoma de las plantas
no traducidas (intrones) sugieren que los viroides evolucionaron a partir de los intrones.
MATRICULA: 2011-0546 GRUPO:5
ResponderEliminarOtros biones acelulares (diap#20)
Satelites(schneider): Un satélite es un agente subvírico formado por ácido nucleico y que depende de la coinfección de la célula huésped por un virus (helper o máster). Si el satélite codifica una proteína de cubierta en la que se introduce el ácido nucleico, hablamos de virus satélite. Un ejemplo es el virus satélite del mosaico del tabaco, que utiliza al virus del mosaico del tabaco para llevar a cabo su reproducción
- virusoides o sateloides(fransky): aunque son estudiados en virología, no se les considera verdaderos virus sino partículas subvirales, por necesitar de otros virus para propagarse. Por esto, en las clasificaciones de virus aparecen con el término de "ARN satélite circular".
- Los plásmidos : son moléculas de ADN extracromosómico circular o lineal que se replican y transcriben independientes del ADN cromosómico. Están presentes normalmente en bacterias, y en algunas ocasiones en organismos eucariotas como las levaduras. Su tamaño varía desde 1 a 250 kb. El número de plásmidos puede variar, dependiendo de su tipo, desde una sola copia hasta algunos cientos por célula.
Priones I (diap#21):· Los priones son partículas infecciosas proteicas que provocan las encefalopatías espongiformes transmisibles.
· Poseen la misma secuencia aminoacídica de una proteína normal, pero presentan una estructura diferente.
· Inducen, por un mecanismo hasta ahora desconocido, la transformación de proteínas normales en anómalas
Priones II (diap#22):
Contradicen el dogma central de la biología molecular, según el cual el flujo de información es en todos los seres vivos: ADN ‡ ARNm ‡ Secuencia Aminoácidos ‡ Estructura Tridimensional Proteínas. La teoría de los priones propuesta por Prusiner supone la existencia de dos plegamientos para una única secuencia de amino ácidos. Además, el replegamiento de la PrP normal por acción de la PrP patológica, implica un flujo de información de una proteína a otra a nivel de estructura terciaria
DIAPOSITIVA # 1: VIROLOGIA
ResponderEliminar2011-0226
La virología (rama de la microbiologia) es el estudio de los virus: su estructura, clasificación y evolución, su manera de infectar y aprovecharse de las células huésped para la reproducción del virus, su interacción con los organismos huéspedes, su inmunidad, la enfermedad que causan, las técnicas para su aislamiento, cultivo y su uso en investigación y terapia. La virología es considerada un sub-campo de la microbiología y la medicina. La virologia también es una rama taxonómica de la biología para estudiar los virus y todos sus componentes.
Su investigación incluye:
la replicación viral
los patógenos virales.
la inmunología viral.
las vacunas virales.
los métodos de diagnóstico.
la quimioterapia antiviral.
las medidas de control de una infección.
los diferentes signos que manifiestan la presencia de virus.
DIAPOSITIVA # 2: ESTRUCTURA GENERAL DE LOS VIRUS
ResponderEliminar2011-0226
Como pudimos observar,en esta imagen se presenta la estructura general de los virus,dentro de las cuales encontramos las siguientes:
El ácido nucleico en los virus contiene la información específica y el potencial para modificar operaciones en la célula infectada.68 Los ácidos nucleicos son macromoléculas constituidas por cadenas de nucleótidos, los cuales a su vez están constituidos por una base nitrogenada asociada a un azúcar del grupo de las pentosas y a uno o más grupos de fosfatos. Existen cuatro posibles tipos de ácido nucleico viral:69 ADN de cadena sencilla, ADN de cadena doble, ARN de cadena sencilla y ARN de cadena doble. Virus que contienen cualquiera de estos tipos de ácido nucleico pueden ser encontrados tanto entre los fagos como entre los virus que infectan a plantas o animales.
El ADN de algunos bacteriófagos se caracteriza por contener bases raras que substituyen alguna o algunas de las bases normalmente presentes en el ADN.70 El ADN de cadena doble presente en algunos virus, se caracteriza por tener segmentos de cadena sencilla en ambos extremos de la molécula. Debido a que son complementarias las secuencias de nucleótidos presentes en ambos extremos. El ADN naturalmente circular puede ser de cadena sencilla como en el fago ØXI74,70 o de cadena doble, como en el virus SV4O. Existe evidencia de que algunos virus ARN que producen infecciones en vegetales como el limonero y la papa contienen moléculas circulares de ARN.
Los virus presentan una amplia diversidad de formas y tamaños, llamadas «morfologías». Son unas 100 veces más pequeños que las bacterias. La mayoría de los virus estudiados tienen un diámetro de entre 10 y 300 nanómetros. Algunos Filovirus tienen un tamaño total de hasta 1.400 nm, sin embargo, sólo miden unos 80 nm de diámetro. La mayoría de virus no pueden ser observados con un microscopio óptico, de manera que se utilizan microscopios electrónicos de barrido y de transmisión para visualizar partículas víricas.Para aumentar el contraste entre los virus y el trasfondo se utilizan tinciones densas en electrones. Son soluciones de sales de metales pesados como wolframio, que dispersan electrones en las regiones cubiertas por la tinción. Cuando las partículas víricas están cubiertas por la tinción (tinción positiva), oscurecen los detalles finos. La tinción negativa evita este problema, tiñendo únicamente el trasfondo.
Una partícula vírica completa, conocida como virión, consiste en un ácido nucleico rodeado por una capa de protección proteica llamada cápside. Las cápsides están compuestas de subunidades proteicas idénticas llamadas capsómeros. Los virus tienen un «envoltorio lipídico» derivado de la membrana celular del huésped. La cápside está formada por proteínas codificadas por el genoma vírico, y su forma es la base de la distinción morfológica.Las subunidades proteicas codificadas por los virus se autoensamblan para formar una cápside, generalmente necesitando la presencia del genoma viral. Sin embargo, los virus complejos codifican proteínas que contribuyen a la construcción de su cápside. Las proteínas asociadas con los ácidos nucleicos son conocidas como nucleoproteínas, y la asociación de proteínas de la cápside vírica con ácidos nucleicos víricos recibe el nombre de nucleocápside.
CONTINUACION
ResponderEliminarEnvoltura
Algunas especies de virus se envuelven en una forma modificada de una de las membranas celulares, o bien es la membrana externa que rodea una célula huésped infectada, o bien membranas internas como la membrana nuclear o el retículo endoplasmático, consiguiendo así una bicapa lipídica exterior conocida como envoltorio vírico. Esta membrana se rellena de proteínas codificadas por el genoma vírico y el del huésped, la membrana lipídica en sí y todos los carbohidratos presentes son codificados completamente por el huésped. El virus de la gripe y el VIH utilizan esta estrategia. La mayoría de virus envueltos dependen de la envoltura para infectar.
La cápsida vírica o cápside vírica
Es una estructura proteica formada por una serie de monómeros llamados capsómeros. En el interior de esta cápside se encuentra siempre el material genético del virus. Puede estar rodeada por una envoltura. Cada capsómero puede estar constituido por una o varias proteínas distintas. El término nucleocápside se refiere al material genético envuelto en su cápside.
DIAPOSITIVA # 3: SIMETRIA VIRALES
ResponderEliminar2011-0226
Hay cuatro tipos principales de morfología vírica:
Helicoidal
las cápsides helicoidales se componen de un único tipo de capsómero apilado alrededor de un eje central para formar una estructura helicoidal que puede tener una cavidad central o un tubo hueco. Esta formación produce viriones en forma de barra o de hilo, pueden ser cortos y muy rígidos, o largos y muy flexibles. El material genético, normalmente ARN monocatenario, pero a veces ADN monocatenario, queda unido a la hélice proteica por interacciones entre el ácido nucleico con carga negativa y la carga positiva de las proteínas. En general, la longitud de una cápside helicoidal está en relación con la longitud del ácido nucleico que contiene, y el diámetro depende del tamaño y la distribución de los capsómeros. El conocido virus del mosaico del tabaco es un ejemplo de virus helicoidal.
Icosaédrica
la mayoría de virus que infectan los animales son icosaédricos o casi-esféricos con simetría icosaédrica. Un icosaedro regular es la mejor manera de formar una carcasa cerrada a partir de subunidades idénticas. El número mínimo requerido de capsómeros idénticos es doce, cada uno compuesto de cinco subunidades idénticas. Muchos virus, como los rotavirus, tienen más de doce capsómeros y parecen esféricos, manteniendo esta simetría. Los ápices de los capsómeros están rodeados por otros cinco capsómeros y reciben el nombre de pentones. Las caras triangulares de éstos también se componen de otros seis capsómeros y reciben el nombre de hexones.
Complejos
los virus tienen una cápside que no es ni puramente helicoidal, ni puramente icosaédrica, y que puede poseer estructuras adicionales como colas proteicas o una pared exterior compleja. Algunos bacteriófagos (como el Fago T4) tienen una estructura compleja que consiste en un cuerpo icosaédrico unido a una cola helicoidal (esta cola actúa como una jeringa molecular, atacando e inyectando el genoma del virus a la célula huésped), que puede tener una base hexagonal con fibras caudales proteicas que sobresalgan.
Los poxvirus son virus grandes y complejos con una morfología inusual. El genoma vírico está asociado con proteínas dentro de una estructura discal central conocida como nucleoide. El nucleoide está rodeado por una membrana y dos cuerpos laterales de función desconocida. El virus tiene una envoltura exterior con una espesa capa de proteína en la superficie. La partícula en general es ligeramente pleomorfa, con una forma que puede ir de la de un huevo a la de un ladrillo. Mimivirus es uno de los tipos de virus más grandes conocidos, con un diámetro en su cápside de 400 nm. De su superficie se proyectan filamentos proteicos de 100 nm. La cápside, tomada en microscopio electrónico, tiene una forma hexagonal, de manera que probablemente es icosaédrica.
Algunos virus que infectan a las Archaeas tienen estructuras inusuales, que no están relacionadas con ningún otro virus conocido. De igual manera, algunos bacteriófagos pueden tener diferentes estructuras en cuanto a su cola, con formas algo raras con respecto a otros virus.
DIAPOSITIVA #4 : TAMA?O DE LOS VIRUS
ResponderEliminar2011-0226
Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus.
Su pequeño tamaño explica lo tardío del descubrimiento de estos agentes. La primera referencia sobre la existencia de los virus se debe al botánico ruso Dimitri Ivanovski en 1892. Este investigador buscaba el agente causante de la enfermedad denominada mosaico del tabaco, y llegó a la conclusión de que debía tratarse de una toxina o de un organismo más pequeño que las bacterias, pues el agente atravesaba los filtros que retenían las bacterias. Denominó a estos agentes patógenos virus filtrables.
En 1897, el microbiólogo holandés Martinus Beijerink realizó experimentos similares a los de Ivanovski, y llegó a desechar la idea de las toxinas, pues se trataba de un agente capaz de reproducirse, ya que mantenía su poder infeccioso de unas plantas a otras, sin diluirse su poder patógeno. Poco después, los microbiólogos alemanes Frederick Loeffler y Paul Frosch descubrieron que la fiebre aftosa del ganado era producida por un virus filtrable que actuaba como un agente infeccioso.
En la década de los 30, con el uso de filtros de tamaño de poro inferior, con las técnicas de cultivo celular in vitro que permitían la obtención de gran cantidad de virus, con la ultracentrifugación y finalmente con el microscopio electrónico y la difracción de rayos X, se logró visualizar a estos agentes.
DIAPOSITIVA #5: FAMILIAS DE VIRUS
ResponderEliminar2011-0226
En esta imagen podemos observar las familias de virus patogenos para el hombre,dentro de los cuales podemos desctacar:
- Bunyavirus:
Género de virus ARN de vertebrados de la familia Bunyaviridae.
-Papovavirus:
Grupo de pequeños virus ADN, algunos de los cuales pueden ser potencialmente productores de cáncer. La verruga humana está producida por un tipo de papovavirus. Algunos tipos de papovavirus son: papovavirus del papiloma, papovavirus del polioma y papovavirus SV.
- Paramixovirus:
Miembro de una familia de virus que abarca a los organismos causantes de la gripe, de las paperas y de algunas infecciones respiratorias.
- Retrovirus:
Virus de ácido ribonucleico que posee una enzima que le permite convertir su material genético en ácido desoxirribonucleico cuando ha penetrado en una célula viva. Al grupo de los retrovirus pertenece el virus del sida.
- Filovirus:
Género de virus con ácido ribonucleico de cadena simple con sentido negativo, de la familia Filoviridae. El género incluye los virus Ébola y Marburg.
- Poxvirus:
Miembro de una familia de virus que incluye a los organismos que producen el molusco contagioso, la viruela y la vacuna.
- Reovirus:
Grupo de tres virus de doble cadena de ARN presentes en todas partes, identificados en los aparatos respiratorio y digestivo tanto de personas sanas como de enfermas. Los reovirus han sido implicados en algunos casos de enfermedades de vías respiratorias superiores y de gastroenteritis infantiles.
- Los Rhabdovirus:
Son virus con cápside helicoidal. Incluye al virus de la rabia y de la estomatitis vesicular (que es el organismo modelo). El virus de la estomatitis vesicular es un virus de 180x75nm, con cápsida rodeada por una envoltura con proyecciones constituídas por la proteína G.
- Togavirus:
Familia de arbovirus a la que pertenecen los organismos causantes de encefalitis, del dengue, de la fiebre amarilla y de la rubéola.
- Coronavirus:
Género de virus de la familia de los Coronaviridae El coronavirus humano es el responsable de infecciones respiratorias de las vías superiores. Miembro de la familia de los virus que abarcan distintos tipos capaces de producir una enfermedad respiratoria aguda.
- Los Calicivirus:
Son Virus RNA pequeños pertenecientes a la familia Caliciviridae. Dentro de esta familia destacan dos géneros: los Norovirus, también conocidos como "virus tipo Norwalk" o "virus pequeños redondos estructurados" y los Sapporovirus.
- Hepadnavirus:
Los hepadnavirus son un subgrupo de los virus que producen hepatitis.
Es uno de los virus más importantes desde el punto de vista médico. Infecta a más de un cuarto de billón de personas y es una de las principales causas de cirrosis y cáncer hepático (carcinoma hepatocelular). HBV, también representa el primer ejemplo del éxito de una vacuna recombinante para una enfermedad contagiosa humana, y puede tener importantes implicaciones para otras enfermedades de transmisión sexual, como el HIV.
-PicoRNAvirus:
Por picornavirus se conoce a todo virus que pertenece a la familia Picornaviridae. Los picornavirus son virus no envueltos, de única cadena ARN con cápside icosaédrica.
Su genoma es inusual ya que tiene una proteína en el extremo 5 ‘que se utiliza como cebador para la transcripción por la ARN polimerasa. El nombre se deriva de “pico”, que significa pequeño, ya que es literalmente un pequeño virus de ARN.
Los picornavirus se separan en una serie de géneros e incluyen muchos importantes patógenos de humanos.
Las enfermedades que causan son muy variadas, que van desde aguda “gripe común” a enfermedades como la poliomielitis, y otras infecciones crónicas.
DIAPOSITIVA #6: DIAGNOSTICO DE LA VIROSIS
ResponderEliminar2011-0226
Los métodos utilizados para reconocer las infecciones por virus humanos pueden clasificarse en DIRECTOS e INDIRECTOS, según persigan demostrar la presencia del virus o de alguno de sus constituyentes (antígeno o genoma viral) o bien la respuesta de anticuerpos específicos por parte del huésped en el curso de la infección.
Gran parte de las técnicas utilizadas en el diagnóstico clínico se basan en pruebas serológicas que identifican anticuerpos específicos frente a diversas proteínas antigénicas. Sin embargo, existen circunstancias en las cuales son necesarias pruebas que detecten precozmente la infección viral (tratamientos específicos, medidas profilácticas, etc.).
En algunas infecciones virales es posible detectar la presencia de antígenos virales previamente al desarrollo de la seroconversión, siendo esta prueba la única evidencia de la exposición al virus cuando no existe aumento de los niveles de anticuerpos circulantes (pacientes inmunodeprimidos).
Igualmente la detección del genoma viral puede favorecer la precocidad del diagnóstico viral y su confirmación. En la última década se han desarrollado una serie de técnicas para el diagnóstico viral basadas en la detección de ácidos nucleicos. De ellas la Reacción en cadena de la polimerasa (PCR) es la más utilizada.
En el momento actual, la tendencia en el diagnóstico virológico consiste en emplear nuevas y más sensibles tecnologías de detección de antígenos y de investigación de ácidos nucleicos con el propósito de lograr un diagnóstico viral más rápido. El desarrollo de quimioterapia antiviral efectiva es responsable de esta tendencia que hace de la identificación rápida, sensible y específica de los virus, una necesidad.
PRINCIPALES TÉCNICAS
METODOS DIRECTOS
Son aquellos que detectan:
1. El virus como agente infeccioso (aislamiento viral).
2. La presencia de antígenos virales (técnicas inmunológicas): Inmunofluorescencia (IF), Enzimoinmunoanálisis (EIA), Test de Aglutinación.
3.La presencia de ácidos nucleicos virales (PCR, etc.).
4.El virus como partícula viral (microscopía electrónica).
DIAPOSITIVA# 7 : ETAPAS DE LA REPLICACION VIRAL
ResponderEliminar2011-0226
En lo que respecta a este tema podemos ver que los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
DIAPOSITIVA#8 : ADHESION
2011-0226
La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado.
DIAPOSITIVA#9: PENETRANCIA
ResponderEliminar2011-0226
Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre las proteínas de la cápsida. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica.
Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras:
Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma.
Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma.
Los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos:
Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma.
Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
DIAPOSITIVA #10: PENETRANCIA DE LOS BACTERIOGRAFOS
2011-0226
El bacteriófago está formado por un corpúsculo que contiene ácido nucleico y una cola con la extremidad en placa mediante la que se fija sobre la bacteria y la infecta. Los bacteriófagos tienen la propiedad de lisar ciertos microbios (colibacilo, bacilos tífico y paratífico, bacilo disentérico, estafilococo, etc) en los que penetra y se multiplica: éstos son los bacteriófagos virulentos que desencadenan cada vez la lisis bacteriana. Los bacteriófagos atenuados pueden, bajo forma de profagos, permanecer inactivos dentro de las bacterias que infectan durante largo tiempo (bacterias lisógenas).
Los bacteriófagos tienen la modalidad de adsorber y penetrar, mejor conocida por su mecanismo de inyección. Las proteínas que forman las espinas de la placa basal entran en contacto mediante sus receptores con los de la pared bacteriana y se anclan a ella.
La accion de algunas enzimas hidroliticas de la capside rompe la pared y esta es perforada por el eje tubular.
El genoma se introduce por el eje hasta el citoplasma de la bacteria quedando fuera de la celula la capsida. Los virus vegetales pasan la pared celular a través de pequeñas grietas producidas por picaduras de insectos.
Los virus animales tienen acceso mas directo a la membrana de las celulas. Los virus sin envoltura penetran por completo en la célula por endocitosis y una vez dentro de la vesicula,el virus libera enzimas hidroliticas para romper la vesicula, y quedar libre en el citoplasma el virus.
Los virus con envoltura penetran al fusionar su cubierta con la membrana celular,y se forma una vesicula endocítica que se une a un endosoma,cuya membrana se fusiona con la envoltura del virus,escapando de ser digerido en su interior
DIAPOSITIVA#11: REPLICACION Y MADURACION
ResponderEliminar2011-0226
La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínas víricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores. Los ribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada.
Los virus con ADN realizan la replicación del material genético de la misma manera que las células; en el caso de los virus con ADN monocatenario, previamente a la replicación se sintetiza una cadena de ADN complementario para formar la doble hélice.
Los virus con ARN replican el material genético sin necesidad de pasar por ADN, actuando cada cadena de ARN como molde para la síntesis de su complementaria.
Los retrovirus constituyen una excepción a lo dicho anteriormente, ya que su ARN sintetiza un ADN bicatenario, que será el que posteriormente realice la síntesis de nuevos ejemplares de RNA vírico.
Los virus con ADN y los retrovirus sintetizan el ARN a partir de la cadena molde de ADN de forma similar a como lo hacen las células.
Los virus con ARN, excepto los retrovirus, sintetiza en el ARN copiando la cadena molde de ARN, sin necesidad de pasar por ADN.
Posteriormente a estos procesos, tiene lugar el ensamblaje de las piezas para construir nuevos viriones. En muchos virus, como el del mosaico del tabaco, el ensamblaje es automático y depende de la concentración salina del medio. En otros virus, en el ensamblaje intervienen enzimas codificadas en el ácido nucleico del virus.
DIAPOSITIVA #12: MADURACION Y SALIDA DE VIRUS DESNUDOS Y ENVUELTOS
ResponderEliminar2011-0226
En esta diapositiva podemos ver un esquema muy detallado de la maduracion y salida de virus desnudos y envueltos.
El desnudamiento ocurre simultáneamente o después de la penetración y comprende la eliminación de todas o parte de las cubiertas proteicas que rodean al ácido nucleico viral, de manera de dejar expuesto dentro de la célula el genoma viral desnudo o asociado a alguna enzima para que pueda expresarse.
En algunos casos sólo se remueve parte de la cápside viral y en esas condiciones el genoma expresa sus funciones. Para otros virus complejos, como por ejemplo los Poxvirus, el desnudamiento parece ocurrir en dos etapas: en la primera hay remoción de ciertas cubiertas proteicas por enzimas celulares, parte del genoma es tanscripto en esas condiciones y luego, en una segunda etapa, y a través de una proteína codificada por el virus, se produce el desnudamiento total del genoma.
En general el desnudamiento es un proceso difícil de estudiar y establecer con precisión sus características por la rapidez con que ocurre y la dificultad en separarlo temporalmente de la penetración. Es por ello que se suele denominar internalización del virión al conjunto de ambos eventos, penetración y desnudamiento.
DIAPOSITIVA#13: SALIDA DE UN VIRUS ENVUELTO
2011-0226
Aqui podemos ver claramente la forma en que sale un virus envuelto;los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes.
No todas las partículas víricas liberadas son infecciosas.
La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
Los virus envueltos (como el VIH) son liberados de la célula huésped por gemación. Durante este proceso, el virus adquiere su envoltura, que es una parte modificada de la membrana plasmática del huésped.
DIAPOSITIVA#14: ANTIVIRALES
ResponderEliminar2011-0226
Los antivirales son un tipo de medicamento usado para el tratamiento de infecciones producidas por virus. Tal como los antibióticos (específicos para bacteria), existen antivirales específicos para distintos tipos de virus. No sin excepciones, son relativamente inocuos para el huésped, por lo que su aplicación es relativamente segura. Deben distinguirse de los viricidas, que son compuestos químicos que destruyen las partículas virales presentes en el medio ambiente.
Muchos de los antivirales disponibles actualmente están diseñados para ayudar el tratamiento del VIH (virus del sida), herpesvirus, productores de la varicela, el herpes labial, el herpes genital, etc. y los virus de la hepatitis B y C, que pueden causar cáncer de hígado. Los investigadores están trabajando actualmente para extender el rango de antivirales a otras familias de patógenos.
Para que un fármaco antiviral sea considerado para el tratamiento de una infección viral debe cumplir ciertos requisitos indispensables. El fármaco debe ser específicamente activo contra el virus “target” (blanco), inhibiendo algún paso esencial de su metabolismo y debe poder debilitar a las cepas resistentes que puedan surgir. Además, debe tener ciertos parámetros que permitan al fármaco ser de alta biodisponibilidad oral, rápida penetración en los tejidos infectados y atóxicos para las células normales y el organismo en general a corto y largo plazo. El fármaco debe ser transformado a su forma activa solamente en células infectadas, debe ser excretado sin ser metabolizado o sin generar metabolitos tóxicos, y debe tener baja frecuencia de administración. Finalmente, el fármaco debe tener bajo costo para que pueda estar disponible para toda la población. Todas estas características, entre otras, dificultan el descubrimiento de nuevos fármacos efectivos para el tratamiento de las enfermedades virales.
DIAPOSITIVA #15: MECANISMOS DE ACCION DE LOS ANTIVIRALES
2011-0226
Los fármacos antirretrovirales no matan al virus de la inmunodeficiencia humana (VIH). Ni siquiera logran dormirlo. Lo 'atontan' un poco y lo mantienen en un estado de reposo, pero que le permite, de vez en cuando, salir a 'pasear' por las células del organismo humano. Lo ha descubierto un equipo de investigadores españoles, que pone así fin a una de las controversias médicas más extendidas en el campo del sida.
Durante años, la teoría más apoyada ha sido la que consideraba que el cóctel de fármacos impedía la replicación del virus. No era capaz de eliminarlo, pero sí podía evitar que siguiera infectando. Otra hipótesis, más minoritaria, indicaba que a pesar de la terapia, el virus podía mantener algo de actividad. Y es esta última la que finalmente se ha confirmado, según la investigación que publica 'Nature Medicine' y que ha sido realizada por expertos del Instituto de Investigación del Sida IrsiCaixa, ubicado en el Hospital German Trias i Pujol de Badalona.
Lo que han visto los investigadores, dirigidos por Javier Martínez Picado, es que algunos pacientes infectados por el VIH, a pesar de que presentan una carga viral indetectable gracias al tratamiento y que parecen tener la infección controlada, cuentan con una pequeña proporción del virus que sigue siendo infeccioso. Un descubrimiento que, según sus autores, "tiene importantes implicaciones clínicas, ya que podría explicar por qué, a pesar de que los pacientes permanecen en terapia durante muchos años, los fármacos no permiten curar la infección de manera definitiva".
Martínez Picado lo aclara de forma sencilla a ELMUNDO.es: "Un paciente infectado sin tratamiento es como un bosque en llamas. Con los fármacos logramos apagar el fuego, pero quedan rescoldos y basta con un pequeño golpe de aire para que vuelva a incendiar. Esto es lo que hace el virus. Está apagado, pero no muerto".
DIAPOSITIVA#16 : RETROVIRALES
ResponderEliminar2011-0226
Dentro de los retrovirales podemos encontrar los siguientes grupos:
-Inhibidores nucleosídicos de la transcriptasa inversa. Actúan sobre la enzima transcriptasa inversa, incorporándola a la cadena de ADN que crea el virus. Con ello, esa cadena se vuelve defectuosa e impiden que el virus se reproduzca.
Son los siguientes: Zidovudina, abacavir, didanosina, estavudina, lamivudina y tenofovir.
-Inhibidores no nucleósidos de la transcriptasa inversa. Bloquean directamente la acción de la enzima y la multiplicación del virus.
Son los siguientes: efavirenz, nevirapina y etravirina.
-Inhibidores de la proteasa. Actúan sobre la enzima proteasa, bloqueando su acción e impidiendo la producción de nuevas copias de células infectadas por el VIH.
Son los siguientes: amprenavir, atazanavir, darunavir, indinavir, lopinavir/r, nelfinavir, ritonavir y saquinavir.
-Inhibidores de la fusión. Impiden la entrada de virus a la célula, con lo que no puede reproducirse.
La enfuvirtida pertenece a este grupo.
-Inhibidores de la integrasa. Bloquean la actividad de la enzima integrasa, responsable de la inserción del ADN del VIH en el ADN humano (código genético de la célula). De esta manera inhibe la replicación del virus y su capacidad de infectar a nuevas células.
DIAPOSITIVA#17,18 Y 19: BIONES ACELULARES
2011-0226
Aqui podemos encontrar a los viroides 1 y 2.
- Pequeñas moléculas de ARN circular de cadena sencilla formadas por unos pocos cientos de nucleótidos. Carecen de
cápsida y su tamaño es una milésima parte del de los virus más pequeños.
- Aparecen con mayor abundancia en el núcleo de las células infectadas.
- Sólo han sido detectados en plantas, en las que producen una gran variedad de enfermedades y pueden ser transmitidos tanto horizontal como verticalmente.
- No existen ninguna evidencia de que los viroides son traducidos a proteínas, ni se conoce cómo causan las enfermedades. Lo que sí se sabe es que son replicados por los enzimas del huésped.
- Algunas semejanzas entre la secuencia de nucleótidos de los viroides y ciertos secuencias del genoma de las plantas
no traducidas (intrones) sugieren que los viroides evolucionaron a partir de los intrones.
DIAPOSITIVA #20: OTROS BIONES ACELULARES
ResponderEliminar2011-0226
AQUI PODEMOS ENCONTRAR LO SIGUIENTE:
-Satelites(schneider): Un satélite es un agente subvírico formado por ácido nucleico y que depende de la coinfección de la célula huésped por un virus (helper o máster). Si el satélite codifica una proteína de cubierta en la que se introduce el ácido nucleico, hablamos de virus satélite. Aunque su genoma no guarda relación con el del helper, sí que depende de las enzimas de éste para replicarse.
Un ejemplo es el virus satélite del mosaico del tabaco, que utiliza al virus del mosaico del tabaco para llevar a cabo su reproducción
- virusoides o sateloides(fransky): aunque son estudiados en virología, no se les considera verdaderos virus sino partículas subvirales, por necesitar de otros virus para propagarse. Por esto, en las clasificaciones de virus aparecen con el término de "ARN satélite circular". De hecho, el término virusoide ha sido también usado genéricamente para referirse a todos los ácidos nucleicos satélites.
- Los plásmidos : son moléculas de ADN extracromosómico circular o lineal que se replican y transcriben independientes del ADN cromosómico. Están presentes normalmente en bacterias, y en algunas ocasiones en organismos eucariotas como las levaduras. Su tamaño varía desde 1 a 250 kb. El número de plásmidos puede variar, dependiendo de su tipo, desde una sola copia hasta algunos cientos por célula.
DIAPOSITIVA #21: PRIONES
2011-0226
Un prion1 es un tipo de proteína patógena (es decir, una molécula compleja que induce enfermedades), más específicamente una psialoproteína patógena, la cual tiene alterada su estructura secundaria, teniendo un incorrecto plegamiento de su estructura terciaria.2 A diferencia del resto de los agentes infecciosos (virus, bacterias, hongos etc...), que contienen ácidos nucleicos (ya sea ADN, el ARN, o ambos), un prion sólo esta compuesto por aminoácidos y no presenta material genético.
El prion, palabra acuñada en 1982 por Stanley B. Prusiner al investigar una serie de enfermedades de carácter crónico e irreversibles que afectaban al sistema nervioso central,3 es un acrónimo inglés derivado de las palabras proteína e infección. Los priones son los responsables de las encefalopatías espongiformes transmisibles en una variedad de mamíferos, incluida la encefalopatía espongiforme bovina (EEB, también conocida como "enfermedad de las vacas locas") en el ganado y la enfermedad de Creutzfeldt-Jakob (ECJ) en humanos.4 Dichas proteínas mutadas forman agregados supramoleculares y son patógenas con plegamientos anómalos ricos en láminas beta, y autoreproducibles.
Los priones se propagan mediante la transmisión de proteínas anómalas con mal plegamiento. Cuando un prion entra en un organismo sano, actúa sobre la forma normal del mismo tipo de proteína existente en el organismo, modificándola y convirtiéndola en prion. Estos priones recién formados pueden pasar a convertir más proteínas, provocando una reacción en cadena que produce grandes cantidades de la proteína prion. Todos los priones conocidos inducen la formación de amiloides plegado, en los que actúan polimerasas formando un agregado que consiste en apretadas hojas ß. El período de incubación de las enfermedades priónicas se determina por la tasa de crecimiento exponencial asociados con la replicación de priones, que es un equilibrio entre el crecimiento lineal y la rotura de los agregados (hay que tener en cuenta que la propagación del prion depende de la presencia de la proteína normalmente plegada en la que los priones pueden inducir plegamiento. Los organismos que no expresan la forma normal de la proteína priónica no pueden desarrollar o transmitir la enfermedad5 .
ResponderEliminarDIAPOSITIVA#22: PRIONES II
2011-0226
No se conocen con exactitud los mecanismos a través de los cuales la Prpc pasa a ser PrpSc. Una de las hipótesis es la rotura de los puentes de disulfuro entre aminoácidos de la estructura de hélice alfa (normalmente entre cisteínas que es un tipo de aminoácido que puede formar dichos puentes), hecho que da lugar a radicales –SH, presentes en las láminas beta.
Según las hipótesis actuales, los aminoácidos que actúan en el cambio conformacional son la metionina, por su capacidad de cambiar el radical metilo por –H consiguiendo un radical -SH; la cisteína, por su disposición a formar puentes disulfuro; y la valina, por su proximidad atómica a la Met y Cys.
En el laboratorio podemos romper los puentes disulfuro mediante dos mecanismos: a partir de la oxidación con ácido perfórmico o mediante la rotura con beta-mercaptoetanol y posterior acetilación con iodoacetato.
Otras hipótesis señalan que la sustitución del aminoácido prolina por leucina puede ser el posible responsable de la desestabilización de la estructura terciaria de la proteína y su transformación en agente patógeno, hecho apoyado con la comparación del gen PRNP procendente de un paciente enfermo con el de un paciente sano.
Mat:2011-0054
Eliminar#1 Virología
Virología es el estudio de virus y virus como agentes: clasificación y evolución, su estructura, sus formas de infectar y explotar las células para la reproducción del virus, las enfermedades que causan, las técnicas para aislar y la cultura de ellos y su uso en la investigación y terapia.Virología es considerada una parte de la microbiología o de patología.
Una importante rama de la Virología es la clasificación de virus. Virus pueden clasificarse según la célula huésped que infectan: virus animales, virus de plantas, hongos virus y bacteriófagos (virus que infectan bacterias, que son los virus más complejos).
Otra clasificación utiliza la forma geométrica de su cápside (a menudo una hélice o un icosaedro) o la estructura del virus (por ejemplo, la presencia o la ausencia de un envolvente de lípidos). Gama de virus en tamaño de unos 30 nm a aproximadamente 450 nm, lo que significa que la mayoría de ellos no puede verse con microscopios de luces.
mat:2011-0054
Eliminar#2 Estructura general de los virus
Los virus más pequeños y simples están constituidos únicamente por ácido nucleico y proteínas. El ácido nucleico es el genoma viral, ubicado en el interior de la partícula, y puede ser ADN o ARN. Generalmente está asociado con un número pequeño de moléculas proteicas que pueden tener actividad enzimática o cumplir alguna función estabilizadora para el plegamiento del ácido nucleico y armado de la partícula viral. Este conjunto de genoma y proteínas íntimamente asociadas es llamado "core", núcleo, nucleoproteína, onucleoide. Este núcleo central está rodeado por una cubierta proteica, la cápside, que junto con el genoma constituye la nucleocápside. Las cápsides virales están formadas por un gran número de subunidades polipeptídicas que se ensamblan adoptando una simetría de tipo helicoidal (nucleocápside en forma de bastón) o icosaédrica (partículas casi esféricas). En algunos virus más complejos, por fuera de la cápside se encuentra otra cubierta, la envoltura, que es una estructura membranosa constituida por lípidos y glicoproteínas. Dicha cubierta viral puede ser considerada una cubierta protectora adicional.
Los virus se caracterizan, a diferencia de los otros organismos, por presentar una única especie de ácido nucleico constitutiva que puede ser ADN o ARN, monocatenario o bicatenario con estructura de doble hélice.
La cápside es una cubierta proteica externa que encierra y protege al genoma viral de la acción de nucleasas y otros factores adversos del medio exterior. Además, en los virus desnudos carentes de envoltura, la cápside es la encargada de establecer a través de alguna de sus proteínas la unión con la célula que será parasitada por el virus. Asimismo, las proteínas de la cápside contienen los determinantes antigénicos contra los que el sistema inmune del huésped elaborará la respuesta de anticuerpos en defensa del organismo.
La envoltura de un virus es una membrana constituida por una doble capa lipídica asociada a glicoproteínas que pueden proyectarse en forma de espículas desde la superficie de la partícula viral hacia el exterior.
Los virus adquieren su estructura mediante un proceso de brotación a través de alguna membrana celular. El número de glicoproteínas que presentan los virus animales es muy variable.
mat:2011-0054
Eliminar#3 Simetría virales
El conocimiento de la estructura viral es necesario para comprender la interacción de las partículas virales con los receptores de la superficie celular y con los anticuerpos neutralizantes. Será la base para idear medicamentos específicos capaces de bloquear la adherencia del virus, el desprendimiento de su envoltura o las dos cosas en las células susceptibles.
La simetría es la forma que adopta un virus en el espacio y está dad por la estructura de su nucleocápside. La simetría puede ser helicoidal, icosaédrica o compleja.
Los virus con simetría helicoidal presentan una nucleocápside cilindrica que puede estar extendida como en el virus del mosaico del tabaco. Este es un virus desnudo. En otros casos la nucleocápside está arrollada sobre sí misma y recubierta por una envoltura, como en el virusInfluenza. Toda la simetría cúbica observada en los virus de los animales, hasta la fecha, es del patrón icosaédrico, la disposición más eficiente de subunidades en una cubierta cerrada. Conocer las reglas que orientan la simetría icosaédrica hace posible determinar el número de capsómeros en una partícula, característica importante de la clasificación de los virus.
El ácido nucleico viral se condensa dentro de las partículas isoméricas; en la condensación del ácido nucleico en una forma adecuada para su acumulación participan proteínas "nucleares" codificadas por el virus o, en el caso de los papovirus, histonas celulares. No han podido definirse aún las reglas para la incorporación del ácido nucleico en las partículas isométricas; probablemente una "secuencia de empaque" participa en el ensamble, aunque en general no son cruciales las estructuras primarias, secundarias y terciarias del ácido nucleico. Existen limitaciones de tamaño en las moléculas de ácido nucleico que se pueden empacar o acumular en una cápside icosaédrica determinada. Las cápsides icosaédricas se forman de manera independiente del ácido nucleico. La mayor parte de los preparados de virus isométricos contendrán algunas partículas "vacías", desprovistas de ácido nucleico viral. Ejemplos de simetría cúbica son los grupos de virus tanto de ADN como de ARN.
Los virus con simetría icosaédrica son poliedros regulares con 20 caras triangulares, 30 aristas y 12 vértices. Son ejemplos de virus con simetría icosaédrica desnudos el virus de la poliomielitis o losadenovirus. Los herpesvirus y togavirus son virus con simetría icosaédrica y recubiertos por una envoltura. En los casos de simetría helicoidal, las subunidades proteínicas están fijas de manera periódica al ácido nucleico viral, con lo que lo amplían en una espiral. El complejo filamentoso de proteína y ácido nucleico viral (nucleocápside) se enrolla a continuación dentro de una cubierta que contiene lípidos. Por lo tanto, a diferencia del caso de las estructuras icosaédricas, existe una interacción periódica regular entre la proteína de la cápside y el ácido nucleico en el virus que tiene simetría helicoidal. No es posible que se formen partículas helicoidales "vacías".
Todos los ejemplos conocidos de los virus de animales con simetría helicoidal contienen genomas de ARN y, con excepción de los rabdovirus, cuentan con nucleocápsides flexibles que están enrolladas en una esfera dentro de las cubiertas.
Se denominan virus de simetría compleja a aquellos que presentan una nucleocápside helicoidal o icosaédrica recubierta por una envoltura laxa. Ejemplos de éstos virus son los poxvirus (forma de ladrillo), los rhabdovirus (forma de bala) y algunos bacteriófagos que poseen una cabeza con simetría icosaédrica y una cola con simetría helicoidal. Algunas partículas virales no presentan simetría cúbica o helicoidal simple, sino que tienen una estructura más complicada. Por ejemplo, los poxvirus tienen forma de ladrillos con rebordes sobre su superficie exteARN, y un centro o núcleo y cuerpos laterales en su interior
mat:2011-0054
Eliminar#4 Tamaño de los virus
Aquí se explica el tamaño que poseen los diferentes virus, La mayoría de los virus poseen un tamaño muy inferior al de las bacterias. Por ello, sólo pueden observarse con el microscopio electrónico.
Dado su pequeño tamaño es necesario utilizar para su medición una medida inferior al micrón, denominada nanómetro (nm) o milimicrón (mu) que equivales a 10-9 metros.
Los virus mas pequeños, por ejemplo los que integran la familia Picornaviridae, donde se incluye el virus de la poliomielitis, miden alrededor de 27 nm y los más grandes, como los de la familiaPoxviridae, donde se encuentra el virus de la viruela, miden aproximadamente 250 nm. Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus.
mat:2011-0054
Eliminar#5 Familias de virus
Inicialmente después de que los virus fueran descubiertos no había sistema para clasificar virus. Por Lo Tanto los virus fueron nombrados casual. La mayor parte de los virus vertebrados se han nombrado según:
las enfermedades asociadas (virus de la polio, rabia)
el tipo de enfermedad causado (virus murine de la leucemia),
los sitios en el cuerpo afectado o de cuál fue aislado el virus primero (rinovirus, adenovirus)
los lugares de donde primero fueron aislados (virus de Sendai, Coxsackievirus)
los científicos que los descubrieron (virus de Epstein-Barr), o
debido a la influencia cultural común del ` de la gripe de las opiniones e.g' del espíritu maligno malo del ` del aire o de la dengue'
Según la clasificación, los virus se agrupan según sus propiedades, no las células que infectan. Las consideraciones principales eran el tipo de DNA nucléica del ácido o ARN.
Cuatro características debían ser utilizadas para la clasificación de todos los virus:
Tipo del ácido nucléico incluyendo la talla del genoma, el strandedness (simple o doble), el lineal o circular, el positivo o el negativo (sentido), segmentos (número y talla), serie y G+C Etc. contento.
Simetría del shell de la proteína
Presencia o ausencia de una membrana del lípido
Dimensiones o la talla del virion y del capsid
Otras propiedades incluyen las propiedades fisicoquímicas incluyendo la masa molecular, pH, estabilidad térmica, susceptibilidad a las substancias químicas y a los extremos físicos y al éter y a los detersorios.
Según la clasificación, los virus se agrupan según sus propiedades, no las células que infectan. Las consideraciones principales eran el tipo de DNA nucléica del ácido o ARN.
Cuatro características debían ser utilizadas para la clasificación de todos los virus:
Tipo del ácido nucléico incluyendo la talla del genoma, el strandedness (simple o doble), el lineal o circular, el positivo o el negativo (sentido), segmentos (número y talla), serie y G+C Etc. contento.
Simetría del shell de la proteína
Presencia o ausencia de una membrana del lípido
Dimensiones o la talla del virion y del capsid
Otras propiedades incluyen las propiedades fisicoquímicas incluyendo la masa molecular, pH, estabilidad térmica, susceptibilidad a las substancias químicas y a los extremos físicos y al éter y a los detersorios
mat:2011-0054
Eliminar#6 Diagnostico de los viroides
Actualmente se han caracterizado 200 especies de viroides que infectan solamente plantas superiores. Los viroides pueden infectar monocotiledóneas y dicotiledóneas, y tanto especies leñosas como herbáceas. La gama de huéspedes de los viroides es muy variable. Las enfermedades más conocidas causadas por viroides en plantas (y de mayor transcendencia económica) son:Cadang-Cadang del cocotero; Exocortis de los cítricos; Piel de manzana marcada por cicatrices; enfermedad del aguacate tostado/quemado por el sol, el atrofiamiento del tomate, la enfermedad del tubérculo filiforme de la patata y La enfermedad de las patatas ahusadas.
A diferencia de los virus vegetales, los viroides se replican, acumulan y expresan sus síntomas mucho más eficientemente a temperaturas elevadas (30-33 °C) (Sänger y Ramon, 1975) y a intensidades de luz asimismo altas. Los viroides -al igual que los priones- son lo máximo en simplicidad y han sido descubiertos hace relativamente poco tiempo. Constituidos por moléculas de ARN desnudo se encuentran en las plantas y se replican en células susceptibles. La molécula puede ser líneal (dando lugar a una estructura en horquilla) o circular (de forma aplanada); en todo caso, los pares de bases complementarios se unen dando como resultado la estructura de doble cadena, semejante a la del ADN. El primer viroide caracterizado, conocido como PSTVd, es una molécula de ARN de cadena simple que contiene 359 nucleótidos.
Los viroides no causan enfermedades en los humanos ni en los animales; sus efectos se limitan a las plantas. Sin embargo, un agente infeccioso similar, un viruosoide, causa la hepatitis D, una variedad excepcionalmente virulenta de esa enfermedad. Los virusoides son moléculas de ARN de una sola hebra, como los viroides, pero requieren un virus asistente para infectar. En el caso de la hepatitis D, el viruosoide necesita de la cubierta proteica del virus de la hepatitis B para transmitirse.
mat:2011-0054
Eliminar#7 Etapas de la replicación viral
El ciclo de replicación de los virus consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden sobrevivir por si mismos en el exterior. Para reproducirse se introducirán en células donde controlarán sus mecanismos reproductivos.
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
mat:2011-0054
Eliminar#8 Adhesión
El primer paso en la infección viral es la absorción a la membrana de la célula susceptible por medio de la adhesión de ligandos virales (proteínas de la capside o glucoproteínas de las espículas, por ejemplo a receptores superficiales de la célula). La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus. Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor.
La mayoría de los fagos se adsorben (pegan) a la pared celular de las bacterias susceptibles, pero algunos fagos también son capaces de adsorberse a los flagelos, vellosidades (pili) o cápsulas presentes en la superficie de la bacteria hospedera. El caso mejor documentado de adsorción viral está representado por los fagos T2 y T4, los cuales son virus con morfología compleja que incluye una cabeza, cola, placa basal, clavijas y fibras de la cola. La pegada inicial de estos fagos a los receptores presentes en la superficie de la bacteria ocurre por medio de los extremos distales de las fibras de la cola. Estas fibras largas que hacen la primera unión se doblan por en medio, de manera que sus extremos distales hacen contacto con la pared celular a muy corta distancia del punto medio de la partícula viral. Después de ocurrida la adhesión, la partícula viral se aproxima a la superficie de la célula. Cuando la placa basal del fago se encuentra a unos 100Å (angstrom= 10-10m) de la pared celular, se establece contacto entre las pequeñas clavijas de la placa basal y la pared celular, pero no existe evidencia de que la propia placa basal se adhiera a la pared celular
mat:2011-0054
Eliminar#9 Penetracion
Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre lasproteínas de la cápsida. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica.
El caso mejor estudiado de la penetración de un virus en la célula hospedera está representado por el caso del fago T2. La cola de este fago es contráctil y en su forma extendida consiste de 24 anillos de subunidades que forman una funda que rodea a un elemento central. Cada anillo consta de 6 subunidades pequeñas y 6 subunidades mayores. Después de la adsorción del fago a la pared celular, ocurre una contracción de la cola que resulta en una fusión de las subunidades pequeñas y grandes para dar 12 anillos de 12 subunidades. El núcleo de la cola no es contráctil y por lo tanto es expulsado e impulsado a través de las capas externas de la bacteria; a continuación, la cabeza del fago se contrae y esto resulta en la inyección del ADN viral en la célula bacteriana. Este proceso posiblemente es facilitado por la presencia de la enzima lisozima en la cola del fago; esta enzima es capaz de digerir las proteínas de las cubiertas bacterianas; además, hay 144 moléculas de adenosina trifosfato (ATP) en la funda de la cola del fago; la energía para la contracción de esta funda proviene de la conversión de la ATP en adenosina difosfato (ADP) por medio de una reacción hidrolítica que libera un grupo fosfato de la ATP
Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras:
Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma.
Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma.
Los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos:
Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma.
Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
mat:2011-0054
Eliminar#10 Penetración de los bacteriófagos
Muchos bacteriófagos carecen de fundas contráctiles y todavía se desconoce el mecanismo detallado por medio del cual penetran en las bacterias. Sin embargo, existe evidencia de que algunos de estos fagos introducen en la célula material proteico además del ácido nucleico.
Las células animales carecen de pared celular y sólo están rodeadas por la membrana plasmática que es muy flexible y cambiante en sus componentes estructurales. Las células animales continuamente introducen elementos del medio externo por medio del proceso de pinocitosis y a través de un procedimiento similar, pero inverso, exportan al medio diversas substancias como enzimas, hormonas y neurotransmisores. Los virus animales solamente pueden infectar células que poseen receptores específicos para el virus en particular. Por lo tanto, se requieren diferentes receptores para diferentes tipos de virus. Se conoce con poco detalle el mecanismo preciso por medio del cual los virus penetran en las células animales. Buena parte del problema radica en que la mayoría de las partículas virales que infectan a una célula son incapaces de iniciar con éxito la multiplicación del virus.
Los virus con envoltura pueden penetrar a la célula hospedera por medio de la fusión entre las envolturas virales y la membrana de la célula. Tanto los virus con envoltura como los virus que carecen de ésta pueden ser introducidos a la célula por medio del proceso de pinocitosis (figura III.3.). La fusión de membranas ocasiona la liberación del genoma viral en el citoplasma de la célula, pero en el caso de la pinocitosis el virus entero es contenido en una vesícula formada a partir de la membrana plasmática. Es probable que esta vesícula se fusione a su vez con alguna de las membranas internas de la célula y la cubierta viral es modificada a consecuencia de esta fusión, lo que da lugar a la liberación del ácido nucleico viral. Es importante hacer notar que la penetración del virus y la subsecuente eliminación de las envolturas virales no implican forzosamente la presencia intracelular de ácido nucleico viral desnudo. El ácido nucleico viral puede permanecer unido a las proteínas internas del virus, las cuales pueden incluir enzimas necesarias para la replicación del virus. En el caso de cierto tipo de virus ARN en los cuales el genoma viral sirve directamente como ARN mensajero (el ácido nucleico a partir del cual se sintetizan las proteínas), el ARN viral se asocia con los ribosomas de la célula. Esto ejemplifica el hecho de que el ácido nucleico viral nunca permanece extendido como un verdadero filamento dentro de la célúla, sino que se enrolla y asocia con proteínas de manera que alcanza un estado favorable de mínima energía libre.
mat:2011-0054
Eliminar#11 Replicación y Maduración
Cuando se han sintetizado un número crítico de varios componentes virales, éstos se ensamblan en partículas víricas maduras en el núcleo y/o citoplasma de la célula infectada. En este ensamblaje y al contrario de lo que ocurre en los bacteriófagos, no están implicados enzimas biosintéticos especiales, sino que ocurre de forma espontánea debido a la gran afinidad de la cápsida con el ácido nucleico viral. La maduración se puede definir como la fase de la infección viral durante la cual los componentes estructurales del virión se producen y ensamblan con el nuevo ácido nucleico viral para formar la nucleocápsida.
La maduración viral es a veces un proceso ineficaz. Pueden acumularse cantidades excesivas de componentes virales y participar en la formación de cuerpos de inclusión en la célula (ver antes). Como resultado de los profundos efectos dañinos de la replicación viral, se desarrollan por último efectos citopáticos celulares y la célula muere. Sin embargo, hay casos en que el virus no lesiona a la célula y se producen infecciones persistentes y prolongadas
REPLICACION DE VIRUS DNA: Después de la adsorción, penetración y eliminación de la cubierta, el ácido nucleico viral es liberado y migra al núcleo de la célula hospedadora. Posteriormente empieza a transcribirse la porción de DNA vírico correspondiente a los genes tempranos. Tiene lugar la traducción, siendo los productos de estos genes enzimas requeridos para la multiplicación del DNA vírico. En la mayoría de los virus DNA, la transcripción temprana se lleva a cabo con la transcriptasa (RNA polimerasa) del hospedador, salvo en el caso de los poxvirus. Algún tiempo después de la iniciación de la multiplicación del DNA, comienza la transcripción y traducción de los genes tardíos que todavía no habían empezado a expresarse. La síntesis de las proteínas de la cápsida ocurre en el citoplasma de la célula hospedadora. Posteriormente las proteínas de la cápsida migran al núcleo donde tiene lugar la maduración: el DNA vírico y las proteínas de la cápsida se ensamblan para formar las partículas víricas completas.
REPLICACION DE VIRUS RNA: La multiplicación de los virus RNA es esencialmente la misma que la de los virus DNA excepto que los mecanismos para generar el mRNA son distintos en los diferentes grupos de virus RNA. Una vez que el RNA vírico y las proteínas son sintetizados, la maduración tiene lugar de una manera similar en el citoplasma. A diferencia de lo que sucede con el ADN, la replicación del ARN es un fenómeno único de los virus ya que la maquinaria genética de la célula se basa exclusivamente en el uso del ADN como templado (ADN-ADN o ADN-ARN). Por lo tanto, para la duplicación de su genoma, así como acontecía para la síntesis de sus ARNm, los virus ARN deben necesariamente aportar la enzima ARN polimerasa ARN dependiente.
mat:2011-0054
Eliminar#12 Maduración y salida de virus desnudos y envueltos
La liberación de los viriones maduros es el paso final en la multiplicación viral. Los mecanismos de liberación varían según el tipo de virus. En algunos virus la célula hospedadora se desintegra liberando los viriones. También pueden ser liberados a ciertos períodos de tiempo mediante exocitosis o a través de canales especiales (túbulos). El número de partículas víricas que se obtienen varía según el tipo de virus, tipo de célula y las condiciones de crecimiento. El rendimiento medio en plantas y animales va desde varios miles a cerca de un millón de viriones por célula, comparado con los varios cientos de fagos que se obtienen en una célula bacteriana.
En las etapas tardías del ciclo de multiplicación viral las moléculas de ácido nucleico y proteínas virales sintetizadas en la célula infectada deben ensamblarse para formar los nuevos viriones y salir al espacio extracelular para estar en condiciones de infectar nuevas células. Este proceso de maduración y ensamble, también denominado morfogénesis, comprende el armado de la cápside a partir de las unidades estructurales y luego la formación del virión completo. El mecanismo va a diferir en base a dos propiedades del virión: la estructura de la cápside y la presencia o no de envoltura.
El desnudamiento ocurre simultáneamente o después de la penetración y comprende la eliminación de todas o parte de las cubiertas proteicas que rodean al ácido nucleico viral, de manera de dejar expuesto dentro de la célula el genoma viral desnudo o asociado a alguna enzima para que pueda expresarse. En algunos casos sólo se remueve parte de la cápside viral y en esas condiciones el genoma expresa sus funciones. Para otros virus complejos, como por ejemplo los Poxvirus, el desnudamiento parece ocurrir en dos etapas: en la primera hay remoción de ciertas cubiertas proteicas por enzimas celulares, parte del genoma es tanscripto en esas condiciones y luego, en una segunda etapa, y a través de una proteína codificada por el virus, se produce el desnudamiento total del genoma.
En general el desnudamiento es un proceso difícil de estudiar y establecer con precisión sus características por la rapidez con que ocurre y la dificultad en separarlo temporalmente de la penetración. Es por ello que se suele denominar internalización del virión al conjunto de ambos eventos, penetración y desnudamiento.
mat:2011-0054
Eliminar#13 Salida de un virus envuelto
Virus que tiene una envoltura o capa exterior. Esta envoltura proviene de la célula infectada o del huésped, en un proceso llamado “gemación”. Durante el proceso de gemación, las partículas víricas recién formadas se “envuelven” o cubren con una capa externa formada por una pieza pequeña de la membrana plasmática de la célula. La envoltura puede cumplir la función de ayudar a un virus a sobrevivir e infectar a otras células.
Los genomas virales recién sintetizados y los polipéptidos de la cápside se ensamblan para formar los virus descendientes. Las cápsides icosaédricas pueden condensarse en ausencia de ácido nucleico, en tanto que las nucleocápsides de los virus de simetría helicoidal no pueden formarse sin ARN viral. No existen mecanismos especiales para la liberación de virus carentes de cubierta; las células infectadas acaban por experimentar lisis y liberar las partículas virales.
Los virus cubiertos maduran por un proceso de gemación. Se insertan glucoproteínas de cubierta específica del virus en las membranas celulares y, a continuación, las nucleocápsides hacen gemación a través de la membrana a nivel de estos sitios modificados y, al hacerlo así, adquieren su cubierta. La gemación ocurre a menudo a nivel de la membrana plasmática y puede abarcar a otras membranas de la célula. Los virus cubiertos no son infecciosos hasta que han adquirido sus cubiertas. Por lo tanto, no se acumulan de manera típica viriones descendientes infecciosos en el interior de la célula infectada.
Dado que la envoltura está constituida por lipoproteínas, todos los virus con envoltura son muy sensibles a los solventes de lípidos, tales como éter y sales biliares. En la envoltura se encuentran glicoproteínas que constituyen importantes antígenos del virus. Las funciones de la envoltura son similares a las de la cápside. Así, en los virus envueltos, es la envoltura la que presenta las glicoproteínas que permitirán la unión a receptores de la célula huésped. Todos los virus con envoltura son muy sensibles a la desecación, al pH ácido, a los solventes de lípidos y a las sales biliares. Esto hace que su transmisión fecal-oral no sea posible.
En cambio, la mayoría de los virus desnudos son resistentes al medio externo, a la desecación y a los solventes de lípidos. Por esta razón, los virus desnudos pueden penetrar por vía digestiva ya que resisten el pH ácido del estómago. Pueden ser transmitidos por vía fecal.oral, a través de agua o alimentoscontaminados o por manos u objetos. Son ejemplos de virus desnudos el Poliovirus, y losAdenovirus.
mat:2011-0054
Eliminar#14 Antivirales
Los antivirales son un tipo de medicamento usado para el tratamiento de infecciones producidas por virus. Tal como los antibióticos (específicos para bacteria), existen antivirales específicos para distintos tipos de virus. No sin excepciones, son relativamente inocuos para el huésped, por lo que su aplicación es relativamente segura. Deben distinguirse de los viricidas, que son compuestos químicos que destruyen las partículas virales presentes en el medio ambiente.Actualmente se dispone de farmacos antivirales de probada eficacia terapeutica y profilactica para varias infecciones virales comunes y algunas potencialmente mortales. Ademas, continua la busqueda de nuevos farmacos antivirales y nuevas estrategias terapeuticas para el tratamiento de las enfermedades virales.
Farmacos que inhiben procesos especificos del virus, como la union a la celula, la decapsidacion del genoma viral o el ensamblaje de los nuevos virus, o, preferentemente, inhiben la sintesis macromolecular dirigida por el virus. Para que un fármaco antiviral sea considerado para el tratamiento de una infección viral debe cumplir ciertos requisitos indispensables. El fármaco debe ser específicamente activo contra el virus“target” (blanco), inhibiendo algún paso esencial de su metabolismo y debe poder debilitar a las cepas resistentes que puedan surgir. Además, debe tener ciertos parámetros que permitan al fármaco ser de alta biodisponibilidad oral, rápida penetración en los tejidos infectados y atóxicos para las células normales y el organismo en general a corto y largo plazo.
El fármaco debe ser transformado a su forma activa solamente en células infectadas, debe ser excretado sin ser metabolizado o sin generar metabolitos tóxicos, y debe tener baja frecuencia de administración. Finalmente, el fármaco debe tener bajo costo para que pueda estar disponible para toda la población. Todas estas características, entre otras, dificultan el descubrimiento de nuevos fármacos efectivos para el tratamiento de las enfermedades virales.
ANTIVIRALES QUE IMPIDEN LA ENTRADA DEL VIRUS A LA CELULA
Amantadina: actua sobre el virus influenza A. Su efecto principal es inhibir la decapsidacion viral o el desensamblaje del virion durante la endocitosis.
Rimantadina: derivado 4-10 veces mas activa que la amantadina.
Oseltamivir: inhibe la actividad de neuraminidasa del virus influenza.
Zanamivir:inhibe la actividad de neuraminidasa del virus influenza.
Docosanol: interfiere en la fusion del VHS-1 y VHS-2 a las membranas de las celulas del huesped. Es de uso topico.
ANTIVIRALES QUE ACTUAN SOBRE LA FASE DE REPLICACIÓN DEL GENOMA VIRAL
Aciclovir: inhibe la sintesis del ADN viral. Tiene accion sobre los herpesvirus.
Valaciclovir: profarmaco del Aciclovir.
Ganciclovir: inhibe la sintesis del ADN viral. Tiene accion sobre los herpesvirus, pero su caracteristica singular es una potente inhibicion de la replicacion del CMV.
Valganciclovir: profarmaco del Ganciclovir.
Adefovir: Actua como inhibidor competitivo de las polimerasas del ADN y transcriptasas inversas virales con respecto al dATP. Se selectividad se relaciona con una mayor afinidad por la ADN polimerasa del VHB que por las polimerasas de las celulas huesped. Esta aprobado para el tratamiento de la hepatitis B cronica.
Cidofovir: inhibe la sintesis de ADN viral de los herpesvirus.
Penciclovir: inhibe la sintesis de ADN viral de los herpesvirus.
Famciclovir. Profarmaco del Penciclovir de administracion oral.
Fomivirseno. Inhibe la replicacion del CMV humano. Se administra de forma intravitrea.
Foscarnet: actua sobre los herpesvirus y la transcriptasa reversa del HIV. Es inhibidor frente a la mayoria de las cepas de CMV resistentes a ganciclovir y de VHS y VVZ resistentes a aciclovir.
Idoxuridina. Inhibe la sintesis de ADN viral. Actua en especial sobre los herpesvirus y poxvirus.
Ribavirina: se utiliza en el tratamiento de la infeccion por HCV y por el VSR.
Trifluridina.
Vidarabina.
mat:2011-0054
Eliminar#15 Mecanismos de acción de los retrovirales
Los retrovirus son pequeños microorganismos que contienen una única cadena de ARN . Su ciclo vital se basa en la conversión de su ARN geonómico en doble cadena de ADN y posteriormente en la integración de éste último en los cromosomas de la célula huésped en lo que se denomina provirus endógeno. De forma ocasional algunas secuencias celulares son recombinadas con el provirus y a su vez estas recombinaciones pueden transferirse a otra población celular infectada, lo que puede tener importantes consecuencias en las funciones celulares.
Inhibidores de transcriptas inversa: Se trata de fármacos que inhiben la síntesis del ADN complementario a partir del ARN viral, función realizada por la transcriptasa inversa (TI) viral en el interior de la célula infectada. Según el mecanismo de bloqueo de la TI se dividen en:
Inhibidores competitivos (ICTI) análogos de nucleósidos/nucleótidos:Se trata de moléculas de estructura similar a los nucleótidos naturales que, al incorporarse como nucleótido anómalo al centro catalítico de la TI, bloquean y finalizan la síntesis del ADN complementario viral. Para que estos fármacos tengan actividad antiviral es preciso que se fosforilen en el interior de las células humanas, mediante la acción de kinasas celulares.
Inhibidores no competitivos (INCTI) no-análogos de nucleósidos: Estos fármacos se unen a la TI en un lugar alejado del centro catalítico, donde se incorporan los nucleótidos naturales. El mecanismo de inhibición enzimática de estos compuestos se basa en la modificación de la estructura terciaria de la TI, lo que finalmente se traduce en una menor actividad procesiva3. Se han comercializado dos fármacos de esta familia (nevirapina [NVP] y efavirenz [EFV]).
Inhibidores de la integrasa: Una vez sintetizado el ADN complementario, éste pasa al interior del núcleo de la célula para allí integrarse en el ADN celular. La integrasa viral es la enzima responsable de este proceso. Actualmente se encuentran en desarrollo clínico una serie de compuestos que bloquean el ciclo viral al nivel de la integración del ADN complementario en el ADN celular.
Inhibidores de la proteasa: Como se ha señalado, el ADN complementario viral sintetizado por acción de la TI se integra en el ADN de la célula infectada. El ciclo viral continúa cuando el ADN viral setranscribe a ARN mensajero que en los ribosomas celulares se traduce en una poliproteína viral. Esta cadena proteica ha de escindirse en diversos péptidos virales que dan lugar a las proteínas estructurales y funcionales que formarán parte de las nuevas partículas virales. La proteasa viral es la enzima responsable de la hidrólisis de la poliproteína en los diversos péptidos individuales. Existe un grupo de fármacos, los inhibidores de la proteasa (IP), que basan su mecanismo de acción en el bloqueo de esta enzima. De este modo se impide la maduración y ensamblaje de las partículas virales, y la infección de nuevas células diana.
mat:2011-0054
Eliminar#16 Retrovirales
Inhibidores de transcriptasa inversa: La transcriptasa inversa es una enzima viral que en el caso del VIH, este requiere para su reproducción. La inhibición de esta enzima impide el desarrollo de ADN viral basado en su ARN. Existen tres clases:
Inhibidores análogos nucleósidos
Fueron la primera clase de antirretrovirales desarrollados.
Zidovudina: Llamada anteriormente azidotimidina, sus siglas son AZT o ZDV y está comercilaizado como Retrovir® por GlaxoSmithKline (GSK); es un análogo de la timidina
Didanosina: También conocida por su sigla ddI, nombre comercial: Videx®, Videx EC®, análogo de la adenosina.
Zalcitabina: También conocida por su sigla ddC, dideoxicitidina, nombre comercial: Hivid®, análogo de pirimidinaTercer antirretroviral aprobado por la FDA.
Estavudina: También llamada d4T, Nombre comercial: Zerit®, Zerit XR®, análogo de timidina Cuarto antirretroviral aprobado por la FDA el 24 de junio de 1994 y en 1996 para uso pediátrico..
Lamivudina: también llamada 3TC, nombre comercial : Epivir®, análogo de citidina..
Abacavir: también llamado ABC, nombre comercial : Ziagen®, análogo de guanosina.
Emtricitabina: también llamada FTC, nombre comercial : Emtriva®, (anteriormente Coviracil), análogo de citidina,aprobado por la FDA el 2 de julio de 2003.
Festinavir: también llamado BMS-986001. Se trata de un análogo de timidina "de segunda generación" similar a d4T pero sin sus problemas de toxicidad.
EFdA: (4'-etinil-2-fluoro-2'-deoxiadenosina) es un compuesto en desarrollo propiedad de Merck inicialmente desarrollado por la biotecnológica japonesa Yasama Corporation y que ha sido estudiado con el apoyo de amFAR y el NIH de EEUU.
Inhibidores de transcriptasa no nucleósidos
Nevirapina: nombre comercial : Viramune®primero de esta clase aprobado por la FDA el 21 de junio de 1996 en adultos y en 1998 para uso pediátrico. Aprobado en España en 1999. Comercializado por Boehringer-Ingelheim.
Delavirdina: nombre comercial : Rescriptor®aprobado por la FDA el 4 de abril de 1997. Actualmente no es de uso coriente debido a sus efectos secundarios adversos. No aprobado para su uso en Europa.
Efavirenz: nombres comerciales: Sustiva®, y Stocrin®; aprobado por la FDA el 21 de septiembre de 1998.
Etravirina: nombre comercial: Intelence®comercializado en España por Janssen-Cilag. Aprobado en Europa para su uso en pacientes previamente tratados.
Rilpivirina: Nombre comercial Edurant®. Rilpivirina se ha comparado con Efavirenz y he demostrado tener menos efectos secundarios a nivel del sistema nervioso central, pero por otro lado, hubo más fracasos virológicos al tratamiento entre los pacientes que tomaron Rilpivirina frente a los que tomaron Efavirenz, especialmente en los pacientes con cargas virales iniciales de más de 100.000 copias/mL.
MK-1439: Es un nuevo inhibidor de la transcriptasa no nucleósido de Merck, que actualmente está en la fase II/III de desarrollo clinico
mat:2011-0054
Eliminar#16 cont
Inhibidores de proteasa
La proteasa es una enzima requerida por el VIH para el ensamblaje final de los viriones, por lo cual su inhibición resulta en inhibición de la replicación viral.
Saquinavir, comercializado como Invirase® por Roche.
Ritonavir, comercializado como Norvir®, por Abbott. Actualmente se utiliza combinado con todos los inhibidores de proteasa a dosis pequeñas (100 o 200 mg) como potenciador (no como antiviral), ya que es un potente inhibidor del citocromo P450, lo que hace que los niveles en sangre de los inhibidores de proteasa se mantenga más tiempo en cifras óptimas. Es un inhibidor de la proteasa con muchos efectos adversos, pero ha de darse a dosis baja con todos los inhibidores de proteasa para que éstos sean efectivos, la única excpeción es el atazanavir.
Indinavir, comercializado como Crixivan® por Merck.
Nelfinavir, comercializado como Viracept® por Roche.
Amprenavir, nombre comercial: Agenerase® aprobado el 15 de abril de 1999. Fue comercializado por GlaxoSmithKline.Requería de la toma de ocho cápsulas de tamaño muy grande dos veces al día, por lo cual la producción fue finalizada el 31 de diciembre de 2004.
Lopinavir, que éstá comercializado como Kaletra®, si bien incluye también bajas dosis de ritonavir (Kaletra es una combinación a dosis fija de lopinavir y ritonavir). Un inconveniente del Kaletra es que debía conservarse a temperaturas no superiores a 25 ºC, lo que es una dificultad en muchos países en vías de desarrollo.
Atazanavir: nombre comercial : Reyataz® aprobado el 20 de junio de 2003. Comercializado por Bristol-Myers Squibb. Primer inhibidor de la proteasa aprobado para 1 toma al día. Parece estar menos asociado con la lipodistrofia y la dislipidemia. Parece no tener resistencia cruzada con otros inhibidores de proteasa.
Fosamprenavir, nombre comercial: Lexiva® (EEUU) o Telzir® (Europa) pro-fármaco de amprenavir, aprobado el 20 de octubre de 2003. En España está disponible desde 2005. Comercializado por GlaxoSmithKline. En el organismo es metabolizado para formar amprenavir, el cual es el ingrediente activo. Es en realidad una versión de amprenavir de liberación lenta, lo cual reduce la cantidad de comprimidos necesarios. Debe administrarse siempre con ritonavir como potenciador.
Tipranavir o Aptivus® su nombre comercial. Comercializado por Boehringer-Ingelheim. Nuevo inhibidor de la proteasa, que es efectivo contra virus que son resistentes a otros ::inhibidores de la proteasa. Actualmente se utiliza poco. Debe administrarse siempre con ritonavir.
Darunavir o Prezista® su nombre comercial. Efectivo contra virus resistentes a otros inhibidores de proteasa. Se utiliza principalmente como componente de tratamiento de rescate, ya que su elevado precio hace que no sea coste-efectivo utiizarlo para iniciar tratamiento. Debe tomarse siempre potenciado con ritonavir
mat:2011-0054
Eliminar#16 cot.
Inhibidores de transcriptasa análogos de nucleótido
Normalmente, los análogos nucleósidos se convierten en el cuerpo a nucleótidos, por lo cual el saltarse este paso deriva en menor toxicidad y disminuye el tiempo de latencia del fármaco.
Tenofovir disoproxil fumarato: o tenofovir o TDF, nombre comercial: Viread® Aprobado el 26 de octubre de 2001 en EEUU.
tenofovir alafenamida: o TAF. Es un nuevo profármaco de tenofovir, en desarrollo y propiedad de Gilead Sciences. De administración una vez al día. Esta nueva formulcación hace que haya menos tenofovir libre, por lo que se espera que disminuyan los efectos secundarios asociados a TDF, especialmente la toxicidad renal.
CMX 157: es un profármaco de tenofovir en desarrollo y propiedad de Merck. Además de las ventajas de mejor distribución del fármaco activo aportadas por TAF, CMX 157 tiene una farmacocinética que permitiría la administración 1 vez a la semana.
Inhibidores de la integrasa
Medicamentos que evitan la acción de la integrasa, selectivos únicamente al VIH-1. Son potentes antirretrovirales y se ha demostrado cierta eficacia en regímenes sin análogos de nucleósidos, pero siempre en combinación (pej Inhibidor de Proteasa + Inhibidor de integrasa). Los regímenes de tratamiento con inhibidores de integrasa producen una recuperación de CD4 más rápida que otros regímenes. Este hecho, si bien es positivo, no está claro si tiene algún beneficio clínico claro.
Raltegravir o Isentress® su nombre comercial. Se administra vía oral dos pastillas al día. Disponible en España desde 2008. Está en desarrollo una nueva formulación de administración una vez al día.
Elvitegravir , de Gilead Sciences. No está disponible como fármaco independiente. Se administra vía oral una sola pastilla al día. Requiere ser administrado con ritonavir como potenciador, por lo que no estaría libre de los efectos adversos de éste. Se metaboliza por el citocromo P450, por lo que puede tener interferencias medicamentosas con fármacos que se metabolizan por esta vía.
Dolutegravir , de ViiV Healthcare. Actualmente en vías de desarrollo (actualmente en fase II-III). Se administra una vez al día y no requiere ser coadministrado con ritonavir ni con ningún otro potenciador. Se metaboliza por la vía del citorcromo P450, con el consiguiente riesgo de interacciones medicamentosas
mat:2011-0054
Eliminar#17 Biones acelulares
Estos microorganismos se clasifican en biones celulares y estos se reproducen y se divide en procariotas y eucariotas y biones acelulares que se replican, por ejemplo los virus, que tienen ácidos nucleicos y los sin ácidos nucleicos que estan los priones.
El término acelular significa literalmente "sin células". Se aplica principalmente a los virus, pero también a provirus y viroides, que son entidades biológicas no formadas por células, a diferencia de bacterias, compuestas por células procariotas, y de hongos, vegetales y animales, formados por células eucariotas.
mat:2011-0054
Eliminar#18 Viroides 1
Los viroides son agentes infecciosos que, al igual que los virus, tienen un ciclo extracelular que se caracteriza por la inactividad metabólica y unciclo intracelular en el que causan infección al huésped susceptible, pero que a diferencia de los virus, los viroides no poseen proteínas ni lípidos y están constituidos por una cadena cíclica corta de ARN, circular o con forma de varilla, (que no codifica proteínas).1 Es importante decir que tanto su forma intracelular como extracelular son las mismas (ARN desnudo), los mecanismos por los cuales éstos logran causar infección están relacionados con la autocatálisis de su material genético. En sí constituyen una etapa primitiva de los virus. El primer viroide fue descubierto por T. O. Diener en 1978, al intentar identificar el agente causal de una enfermedad que inicialmente supuso era inducida por un virus llamada "la enfermedad del tubérculo fusiforme de la patata" (potato spindle tuber o PSTVd) La nomenclatura de los viroides incluye el sufijo “Vd” para distinguirlos de los virus. El segundo viroide en ser descubierto fue el agente causal de la exocortis de los cítricos (citrus exocortis viroid, CEVd). Actualmente se conocen más de 200 viroides.
mat:2011-0054
Eliminar#19 Viroides 2
Los viroides son los agentes infecciosos de menor complejidad genética y estructural conocidos y representan una forma extrema de parasitismo. Están constituidos exclusivamente por moléculas de ARN de cadena simple, cerradas covalentemente o con forma de bastón de bajo peso molecular (246 a 1994 nucleótidos). Carecen de actividad de ARN mensajero y se replican de forma autónoma, utilizando el sistema de transcripción de la célula suceptible.2 Los viroides se encuentran, casi exclusivamente, en el núcleo de las células infectadas; se desconoce el modo en que se replican pero se sabe que el ARN que los constituyen no funciona como ARN mensajero y tampoco se traduce a enzimas que participen en su propia replicación. Los viroides no codifican para ninguna proteína y llevan a cabo su replicación en la célula hospedadora a través de la RNA polimerasa II codificada por la célula hospedadora aunque también se ha propuesto la posibilidad de que lo haga gracias a una RNA polimerasa dependiente de RNA presente en algunas células eucariotas (Cann, 2001). Dada su localización, se presume que causan la enfermedad interfiriendo con la regulación génica de la célula hospedadora en la etapa de corrección del ARN mensajero celular: particularmente en la eliminación de intrones y de empalmes de exones, presentando secuencias muy similares a los intrones que normalmente son cortados. Sus ARN se replican varias veces.
Actualmente se han caracterizado 200 especies de viroides que infectan solamente plantas superiores. Los viroides pueden infectar monocotiledóneas y dicotiledóneas, y tanto especies leñosas como herbáceas. La gama de huéspedes de los viroides es muy variable. Las enfermedades más conocidas causadas por viroides en plantas (y de mayor transcendencia económica) son:Cadang-Cadang del cocotero; Exocortis de los cítricos; Piel de manzana marcada por cicatrices; enfermedad del aguacate tostado/quemado por el sol, el atrofiamiento del tomate, la enfermedad del tubérculo filiforme de la patata y La enfermedad de las patatas ahusadas.
A diferencia de los virus vegetales, los viroides se replican, acumulan y expresan sus síntomas mucho más eficientemente a temperaturas elevadas (30-33 °C) (Sänger y Ramon, 1975) y a intensidades de luz asimismo altas. Los viroides -al igual que los priones- son lo máximo en simplicidad y han sido descubiertos hace relativamente poco tiempo. Constituidos por moléculas de ARN desnudo se encuentran en las plantas y se replican en células susceptibles. La molécula puede ser líneal (dando lugar a una estructura en horquilla) o circular (de forma aplanada); en todo caso, los pares de bases complementarios se unen dando como resultado la estructura de doble cadena, semejante a la del ADN. El primer viroide caracterizado, conocido como PSTVd, es una molécula de ARN de cadena simple que contiene 359 nucleótidos.
ma:2011-0054
Eliminar#20 Otros biones acelulares
Un virusoide o ARN satélite es una pequeña molécula patógena de ARN (ácido ribonucleico) que se encuentra encapsulada en ciertos virus de plantas con secuencias de ARN de mayor tamaño, lo que se denomina un virus asistente, el cual hospeda al virusoide y es necesario para que éste produzca una infección exitosa.
El ARN circular del virusoide es extremadamente pequeño, solo unos cientos de nucleótidos, pues no necesita genes para nada salvo para replicarse. Los virusoides son similares a los viroides en tamaño y estructura, pero se diferencia en que los viroides no necesitan a un virus asistente.
Aunque son estudiados en virología, no se les considera verdaderos virus sino partículas subvirales, por necesitar de otros virus para propagarse. Por esto, en las clasificaciones de virus aparecen con el término de "ARN satélite circular". De hecho, el término virusoide ha sido también usado genéricamente para referirse a todos los ácidos nucleicos satélites.
Debido a que carecen de estructuras que le faciliten el ingreso a la célula que atacan, para ingresar dependen de un virus, por esto a los virusoides también se les denomina Satélites. Dentro de la célula, el virusoide se adueña de parte del mecanismo de síntesis de la bacteria, y la bacteria comienza a trabajar para el virusoide, formando numerosas réplicas de él, que quedan envueltas junto al material genético de los descendientes del virus que la célula infectada también esta formando. Así, el virus recién formado lleva en su interior una fracción de ARN que no le pertenece, al Satélite, o sea, al virusoide
mat:2011-0054
Eliminar#21 Priones 1
Son agentes patógenos formados por una proteína ( proteína del prión o PPr ). Producen entre otras, la enfermedad de las "vacas locas" o encefalopatía bovina espongiforme. Esta proteína se acumula en el cerebro de animales enfermos , dando lugar a la estructura esponjosa de la corteza cerebral que da nombre a la enfermedad.
Los priones, o las enfermedades producidas por priones, tienen un comportamiento sorprendente, por un lado se transmiten verticalmente , como cualquier enfermedad hereditaria típica, mientras que por otro lado se comportan de manera infectiva, transmitiéndose horizontalmente, mediante contagios que pueden darse entre individuos de distintas especies.
La proteína del prión (PrP) normal, tiene una secuencia de aminoácidos, (estructura primaria) idéntica a la proteína del prión patógena. La diferencia entre las dos recae en la estructuras secundaria y terciaria.
A pesar de que actualmente hay muy poca información sobre la función que tienen las proteínas Prpc (estado no patógeno), algunos experimentos han demostrado que tienen un papel activo en el correcto desarrollo neuronal, que es una proteína capaz de unir específicamente Cu2+ (procesos de oxidoreducción), y también se han relacionado los priones con proteínas de transducción de señales, la adhesión celular y la regulación y distribución de los receptores de acetilcolina.6
mat:2011-0054
Eliminar#22 Priones 2
Una vez obtenido el prion responsable de la patología, comenzó a dilucidarse su estructura. Se trata de una glicoproteína de 27-30 kD, que tiene la misma estructura primaria que una proteína similar presente en el cerebro de la oveja. La modificación estructural, no genética, se debe a un proceso de postraducción. Esto se presenta en funciones muy especiales.
El gen codificando del prion Prpc (gen PRNP) se localiza en el brazo corto del cromosoma 20, no tiene intrones y es un gen autosómico dominante. Se expresa en el tejido neuronal, cardíaco, muscular, pancreático y hepático.
El plegamiento erróneo de la Prpc a PrpSc confiere a la PrpSc dos propiedades que la diferencian de la Prpc: la resistencia parcial a la digestión por proteasas y su insolubilidad. Estas dos propiedades hacen que la PrpSc sea estable y la capacitan para poder formar agregados proteicos responsables de la acumulación de PrpSc en forma de placas amiloides en el tejido nervioso.
Un prion es un tipo de proteína patógena (es decir, una molécula compleja que induce enfermedades), más específicamente una psialoproteína patógena, la cual tiene alterada su estructura secundaria, teniendo un incorrecto plegamiento de su estructura terciaria. A diferencia del resto de los agentes infecciosos (virus, bacterias, hongos etc...), que contienen ácidos nucleicos (ya sea ADN, el ARN, o ambos), un prion sólo esta compuesto por aminoácidos y no presenta material genético.
El prion, palabra acuñada en 1982 por Stanley B. Prusiner al investigar una serie de enfermedades de carácter crónico e irreversibles que afectaban al sistema nervioso central, es un acrónimo inglés derivado de las palabras proteína e infección. Los priones son los responsables de las encefalopatías espongiformes transmisibles en una variedad de mamíferos, incluida la encefalopatía espongiforme bovina (EEB, también conocida como "enfermedad de las vacas locas") en el ganado y la enfermedad de Creutzfeldt-Jakob (ECJ) en humanos. Dichas proteínas mutadas forman agregados supramoleculares y son patógenas con plegamientos anómalos ricos en láminas beta, y autoreproducibles.
Los priones se propagan mediante la transmisión de proteínas anómalas con mal plegamiento. Cuando un prion entra en un organismo sano, actúa sobre la forma normal del mismo tipo de proteína existente en el organismo, modificándola y convirtiéndola en prion. Estos priones recién formados pueden pasar a convertir más proteínas, provocando una reacción en cadena que produce grandes cantidades de la proteína prion. Todos los priones conocidos inducen la formación de amiloides plegado, en los que actúan polimerasas formando un agregado que consiste en apretadas hojas ß. El período de incubación de las enfermedades priónicas se determina por la tasa de crecimiento exponencial asociados con la replicación de priones, que es un equilibrio entre el crecimiento lineal y la rotura de los agregados (hay que tener en cuenta que la propagación del prion depende de la presencia de la proteína normalmente plegada en la que los priones pueden inducir plegamiento. Los organismos que no expresan la forma normal de la proteína priónica no pueden desarrollar o transmitir la enfermedad.
Leomy Anyelina Porro Mercedes Matricula: 2011-0254 Grupo: 05
ResponderEliminarAntes de comenzar hablando de los virus conoceremos que es la
virología
La virología (rama de la microbiologia) es el estudio de los virus: su estructura, clasificación y evolución, su manera de infectar y aprovecharse de las células huésped para la reproducción del virus, su interacción con los organismos huéspedes, su inmunidad, la enfermedad que causan, las técnicas para su aislamiento,
Virus
Un VIRUS BIOLÓGICO se trata de una partícula infecciosa, un organismo que se replica a sí mismo en el interior de las células que ataca (células animales, vegetales o bacterianas).Podemos decir que no es un organismo totalmente vivo, ya que si analizamos la definición de ser vivo nos encontramos con que: nacen, crecen, se reproducen y mueren, de las cuales el virus por si solos solo nacen y mueren, necesitan una célula a la que infectar para crecer y evolucionar. No tienen metabolismo propio, necesitan una célula a la infectar, son “parásitos”.
ESTRUCTURA GENERAL DE LOS VIRUS
En los virus se distinguen las partes siguientes:
1.- Genoma vírico. Se compone de una o varias moléculas de ADN o de ARN, pero nunca los dos simultáneamente, Se trata de una sola cadena, abierta o circular, monocatenaria o bicatenaria. .
2.- Cápsida. Es la cubierta proteica que envuelve al genoma vírico. Está formada por proteínas globulares o capsómeros que se disponen de una manera regular y simétrica, lo que determina la existencia de varios tipos de cápsidas: icosaédricas, helicoidales y complejas.
La función de la cápsida es proteger el genoma vírico y, en los virus carentes de membrana, el reconocimiento de los receptores de membrana de las células a las que el virus parasita.
3.-Envoltura membranosa. Formada por una doble capa de lípidos que procede de las células parasitadas y por glucoproteínas incluidas en ella cuya síntesis está controlada por el genoma vírico. Las glucoproteínas sobresalen ligeramente de la envoltura y tienen como función el reconocimiento de la célula huesped y la inducción de la penetración del virus en ella mediante fagocitosis.
Leomy Anyelina Porro Mercedes Matricula: 2011-0254 Grupo: 05
ResponderEliminarcontinuación:
SIMETRÍA VIRALES
La simetría es la forma que adopta un virus en el espacio y está dado por la estructura de su nucleocápside. La simetría puede ser helicoidal, icosaédrica o compleja.
Los virus con simetría helicoidal presentan una nucleocápside cilindrica que puede estar extendida como en el virus del mosaico del tabaco. Este es un virus desnudo. En otros casos la nucleocápside está arrollada sobre sí misma y recubierta por una envoltura, como en el virusInfluenza.
Los virus con simetría icosaédrica son poliedros regulares con 20 caras triangulares, 30 aristas y 12 vértices. Son ejemplos de virus con simetría icosaédrica desnudos el virus de la poliomielitis o losadenovirus. Los herpesvirus y togavirus son virus con simetría icosaédrica y recubiertos por una envoltura.
Se denominan virus de simetría compleja a aquellos que presentan una nucleocápside helicoidal o icosaédrica recubierta por una envoltura laxa. Ejemplos de éstos virus son los poxvirus (forma de ladrillo), los rhabdovirus (forma de bala) y algunos bacteriófagos que poseen una cabeza con simetría icosaédrica y una cola con simetría helicoidal.
TAMAÑO DE LOS VIRUS
Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus. Algunos filovirus tienen una longitud total de hasta 1400nm.
Leomy Anyelina Porro Mercedes Matricula: 2011-0254 Grupo: 05
ResponderEliminarcontinuación:
FAMILIA DE VIRUS
Hay muchas variedades de familias y de las más importante citaremos:
Familia Parvoviridae
- Virus de pequeño tamaño: son los más pequeños que existen.
- DNA mc lineal
3 géneros:
Gº Dependovirus
Tienen virus defectivos: virus que no tienen capacidad de multiplicarse por sí solos, sino que dependen de otros virus: los adenovirus y los virus herpes.
No provoca infecciones.
Tienen antígeno común de grupo.
Gº Densovirus
Afectan a insectos. Interés: lucha biológica contra plagas. En concreto, afectan a ortópteros, lepidópteros, dípteros.
Multiplicación tipo II de Baltimore, intranuclear.
Exhibe poca información genética: 3-4 genes.
Tiene una cápside proteica, y carece de envoltura de lípidos o glúcidos: es un virus desnudo.
Resistente al éter y al cloroformo.
Muy resistente a altas temperaturas (60º C durante 1 h.) y distintos pH (de 3-9) es muy difícil hacer una desinfección de este género.
Gº Parvovirus
Tienen tropismo por células en división. Provoca enteritis, panleucopenias (leucemias muy graves), depresión del SI.
Gatos: tropismo por el cerebelo. Provoca hipoplasia celebelar (en cachorros, incluso fetos).
Perros: tropismo por el cerebelo y también por el miocardio. Es patógeno para animales jóvenes.
Son virus pantotropos, pero tienen apetencia por los tejidos descritos y el SI:
Rango amplio de hospedadores: perros, gatos, cerdos, gansos, visones.
Familia Papovaviridae
- ADN bc, 1 hebra circular.
- 5-8 genes
- Desnudos resistentes al éter y al cloroformo
- Morfología esférica, simetría cúbica (icosaédrica)
- Estables a distintos pH y a distintas temperaturas (termoestables)
- Mult. Intranuclear, ciclo de multiplicación: tipo I de Baltimore
- Carecen de Ag de grupo, pero tienen antígenos comunes de género.
- Cultivos celulares: efecto lítico y citopático
- Virus transformantes: producen hiperplasias, amontonamientos celulares.
- Cuerpos de inclusión Cowdry A
- Virus hemoaglutinantes
- Transmisión: contacto directo entre individuos y vectores (ácaros, sifonápteros: pulgas)
- Tienen una elevada especificidad por el hospedador
Familia Adenoviridae
“Adeno” significa glándula.
- ADN bc lineal.
- 30 genes
- Morf. esférica simetría cúbica.
- Pequeño tamaño.
- Tiene espículas, y también hemoaglutininas.
- Multiplicación tipo I de Baltimore.
- Tiene proteínas en la cápside, pero no envoltura (ni glúcidos ni lípidos) virus desnudo.
- Resistente al éter y al cloroformo.
- 252 capsómeros.
- No tienen antígeno común de grupo pero sí antígenos comunes de género (interesante para vacunas).
- Estables a distintas temperaturas y pH.
- Efecto citopático, citolítico y también despegamiento celular en las células en cultivo.
- Inducen hiperplasias.
Familia Herpesviridae
Incluidas dentro de la familia hay 3 subfamilias, con distintos géneros. En total hay más de 80 virus que afectan a todas las especies animales: aves, reptiles, anfibios, cerdos, caballos, rumiantes y animales salvajes. También al hombre.
“Herpes” significa reptante. De todos los animales, el menos receptivo a los virus herpes son las ovejas.
- ADN bc lineal
- 160 genes
- Morfología circular, simetría cúbica.
- 162 capsómeros
- Envoltura lipídica y proyecciones
- Sensibles al éter y al cloroformo.
- Partes: ácido nucleico, proteínas (cápside), lípidos y glúcidos.
- Sensibles a la temperatura y al pH (lábiles).
Leomy Anyelina Porro Mercedes Matricula: 2011-0254 Grupo: 05
ResponderEliminarcontinuación
Familia Poxviridae
Comprenden géneros que afectan tanto al hombre como a los animales. Son los virus de las viruelas, los de tamaño más grande que se conocen (visibles al microscopio óptico).
Son virus modélicos, que han servido para el estudio de oncogénesis. Actualmente sirven como vectores vacunales: se les inyecta el ADN exógeno (de virus o de bacteria) y se puede producir lo que sea a partir de ese ADN.
- Morfología y simetría compleja (tienen “cuerpos laterales”, de los que se desconoce la función). El interior de la célula tiene forma oval, y el exterior de ladrillo.
- 300 genes.
- Exhiben proteínas y enzimas.
DIAGNOSTICO DE LAS VIROSIS
Síntomas Los síntomas son muy variables y dependen del virus y del órgano afectado. Muchos virus, como muchas bacterias, producen fiebre y síntomas respiratorios (tos, estornudos) o intestinales (náuseas, vómito, diarrea). Las virosis, aun cuando no sean de peligro, a menudo provocan fiebre elevada en los niños de corta edad.
Diagnóstico Algunas infecciones virales, como la gripe, el resfriado común, la varicela son fáciles de identificar por sus síntomas, sin que se necesiten análisis de laboratorio. Para otras, como la hepatitis viral, el sida y la mononucleosis infecciosa, se suele analizar una muestra de sangre en busca de anticuerpos específicos contra el virus. La presencia de estos anticuerpos confirma el diagnóstico. En ciertos casos, los virus pueden cultivarse en tejidos celulares o bien el virus se identifica por su ácido nucleico, valiéndose de una técnica que recibe el nombre de reacción en cadena de la polimerasa (RCP). Las pruebas como el RCP o el cultivo celular se emplean cuando las de anticuerpos no tienen la precisión suficiente o cuando es necesario cuantificar con mayor exactitud el virus invasor.
ETAPAS DE LA REPLICACION
Adhesión: que depende de la interacción de proteínas de superficie del virus con receptores celulares específicos. Por ejemplo el receptor CD4 del linfocito T y la glicoproteína gp120 del virus HIV, las hemaglutininas de los virus influenza y los receptores de ácido acetil-neuramínico en la superficie de las células epiteliales, etc
Penetración y decapsidación: la penetración a través de la membrana plasmática de la célula puede efectuarse por los siguientes mecanismos: por pasaje directo, por fusión de la membrana celular con la envoltura viral, o por endocitosis o viropexis, mecanismo más frecuente, es un fenómeno similar a la fagocitosis, en el cual el virus es incluído en una vacuola fagocítica. Luego de la penetración se produce la decapsidación, es decir la liberación del ácido nucleico viral de la nucleocápside.
Penetración de los Bacteriófagos
Inyección del material genético viral: Después de la adsorción, se produce un cambio configuracional en las proteínas de la placa basal, alguna de las cuales tienen actividad enzimática y producen un poro en la membrana citoplasmática de la célula. La vaina del fago se contrae y el material genético viral ingresa en la célula, mientras que la envoltura proteica queda en el exterior.
Para entrar en una célula, los fagos se acoplan a receptores específicos en la superficie de la bacteria, que pueden ser lipopolisacáridos, ácidos teicoicos, proteínas o incluso flagelos. Por ello, cada fago solo podrá infectar ciertas bacterias según sus receptores. Puesto que los fagos no son móviles, dependen de encuentros al azar con los receptores adecuados en solución para poder infectar una bacteria.
Parece que los bacteriófagos presentan una especie de jeringa mediante la cual introducen su material genético en el interior de la célula. Tras el reconocimiento del receptor adecuado, la cola y cuello del fago se contraen, quedando así el fago acoplado a la superficie celular. El material genético puede ser ahora introducido a través de la membrana o bien simplemente depositado sobre la superficie. No se descarta que pueda haber fagos con otros métodos diferentes para introducir su material genético en la célula.
Leomy Anyelina Porro Mercedes Matricula: 2011-0254 Grupo: 05
ResponderEliminarcontinuación:
MADURACION Y SALIDA DE VIRUS DESNUDOS Y ENVUELTOS.
El desnudamiento ocurre simultáneamente o después de la penetración y comprende la eliminación de todas o parte de las cubiertas proteicas que rodean al ácido nucleico viral, de manera de dejar expuesto dentro de la célula el genoma viral desnudo o asociado a alguna enzima para que pueda expresarse.
En algunos casos sólo se remueve parte de la cápside viral y en esas condiciones el genoma expresa sus funciones. Para otros virus complejos, como por ejemplo los Poxvirus, el desnudamiento parece ocurrir en dos etapas: en la primera hay remoción de ciertas cubiertas proteicas por enzimas celulares, parte del genoma es tanscripto en esas condiciones y luego, en una segunda etapa, y a través de una proteína codificada por el virus, se produce el desnudamiento total del genoma.
En general el desnudamiento es un proceso difícil de estudiar y establecer con precisión sus características por la rapidez con que ocurre y la dificultad en separarlo temporalmente de la penetración. Es por ello que se suele denominar internalización del virión al conjunto de ambos eventos, penetración y desnudamiento.
SALIDA DE UN VIRUS ENVUELTO.
Vemos aquí como los virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes.
No todas las partículas víricas liberadas son infecciosas.
La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
Los virus envueltos (como el VIH) son liberados de la célula huésped por gemación. Durante este proceso, el virus adquiere su envoltura, que es una parte modificada de la membrana plasmática del huésped.
Maduración: de produce el ensamble de los ácidos nucleicos neoformados con las proteínas de la cápside.
Liberación: los nuevos viriones salen de la célula por lisis de la misma, como el virus polio; o bien por brotación a través de la membrana celular, como los paramixovirus o arenavirus; o de la membrana nuclear, como los herpesvirus o del retículo endoplásmico, como los togavirus.
MECANISMO DE ACCION DE LOS RETROVIRALES
Los antivirales son un tipo de medicamento usado para el tratamiento de infecciones producidas por virus. Tal como los antibióticos (específicos para bacteria), existen antivirales específicos para distintos tipos de virus. No sin excepciones, son relativamente inocuos para el huésped, por lo que su aplicación es relativamente segura. Deben distinguirse de los viricidas, que son compuestos químicos que destruyen las partículas virales presentes en el medio ambiente.
Su eficacia (y a veces su toxicidad) están relacionadas con la competencia inmunológica del paciente.
Su actividad es restringida al actuar sobre un “órgano diana” muy específico.
Desarrollo de Resistencias.
La mayoría solo son eficaces durante la fase de Replicación viral y no sobre los virus Latentes.
La utilización óptima de los antivirales precisa de la realización de un diagnóstico clínico correcto.
Leomy Anyelina Porro Mercedes Matricula: 2011-0254 Grupo: 05
ResponderEliminarcontinuación
RETROVIRUS
Fármacos Antivirales
Las drogas antirretrovirales actúan sobre diferentes pasos necesarios para la replicación viral; hasta la fecha, las más extendidas son aquellas drogas que actúan sobre la transcriptasa inversa y las drogas que actúan sobre las proteasas; aunque ya existen fármacos capaces de inhibir otros sitios tales como los inhibidores de la fusión y los de la integrasa.
Los Inhibidores de la Reversotranscriptasa (IRT) alteran la función de la enzima llamada reversotranscriptasa que es la que utiliza el VIH para cambiar su mensaje genético a una forma que pueda ser fácilmente insertada dentro del núcleo de la célula infectada. Es la encargada de hacer una copia en ADN del ARN viral de manera que éste pueda insertarse en el ADN celular, mediante la acción de las integrasas, y así producir proteínas del VIH necesarias para la auto reproducción viral. Los IRT nucleósidos y los nucleótidos necesitan ser fosforilados para luego interactuar con un sustrato en el sitio de unión a la enzima y, cuando es incorporado, entonces termina la elongación de la cadena de ADN; mientras que los IRT no nucleósidos no necesitan ninguna conversión metabólica y bloquean directamente la reacción con la reverso transcriptasa por una interacción específica con el sitio de unión “sin sustrato” de la reversotranscriptasa del VIH.
Los Inhibidores de las Proteasas (IP) son fármacos con una potente actividad antiviral. La proteasa del VIH es requerida para el clivaje del polipéptido gag-pol en sus partes funcionales y la inhibición de este clivaje trae como consecuencia una disminución en la producción de viriones maduros. O sea, estos medicamentos se parecen a los trozos de la cadena de proteína que la proteasa corta normalmente. Al adherirse a la proteasa, los IP previenen que la proteasa corte las cadenas largas de proteínas y enzimas en trozos más cortos que el virus necesita para reproducirse en nuevas copias de sí mismo. Aunque las cadenas largas no hayan sido cortadas en los trocitos correctos, las nuevas copias del VIH aún se construyen y continúan empujando a través de la membrana de la célula infectada. Pero estas nuevas copias del VIH son “defectuosas”, ya que no han sido formadas completamente, por lo tanto no pueden continuar infectando otras células.(1,2,3,4)
INHIBIDORES DE FUSION
Los inhibidores de la entrada evitan el ingreso del VIH a las células sanas, se adhieren a las proteínas de superficie de las células T o a las proteínas de superficie del VIH. Para que el VIH es una y pueda entrar a las células T, las proteínas de la capa externa del VIH deben unirse a las proteínas de superficie de las células T.
Los inhibidores de la entrada evitan que este proceso ocurra. Algunos inhibidores de la entrada se adhieren a las proteínas gp120 ó gp41 de la capa externa del VIH. Otros inhibidores utilizan la proteína CD4 ó los receptores CCR5 ó CXCR4 de la superficie de las células T. Si los inhibidores de la entrada logran adherirse eficientemente a las proteínas, éstas quedan bloqueadas y entonces el VIH no puede unirse a la superficie de las células T e ingresar a las mismas.
Leomy Anyelina Porro Mercedes Matricula: 2011-0254 Grupo: 05
ResponderEliminarconticuacion:
BIONES ACELULARES
El término acelular significa literalmente "sin células". Se aplica principalmente a los virus, pero también a provirus y viroides, que son entidades biológicas no formadas por células, a diferencia de bacterias, compuestas por células procariotas, y de hongos, vegetales y animales, formados por células eucariotas.
LOS VIROIDES I y II
son agentes infecciosos que, al igual que los virus, tienen un ciclo extracelular que se caracteriza por la inactividad metabólica y unciclo intracelular en el que causan infección al huésped susceptible, pero que a diferencia de los virus, los viroides no poseen proteínas ni lípidos y están constituidos por una cadena cíclica corta de ARN, circular o con forma de varilla, (que no codifica proteínas).1 Es importante decir que tanto su forma intracelular como extracelular son las mismas (ARN desnudo), los mecanismos por los cuales éstos logran causar infección están relacionados con la autocatálisis de su material genético.
Estas pequeñas moléculas de ARN circular de cadena sencilla formadas por unos pocos cientos de nucleótidos. Carecen de cápsida y su tamaño es una milésima parte del de los virus más pequeños.
- Aparecen con mayor abundancia en el núcleo de las células infectadas.
- Sólo han sido detectados en plantas, en las que producen una gran variedad de enfermedades y pueden ser transmitidos tanto horizontal como verticalmente.
- No existen ninguna evidencia de que los viroides son traducidos a proteínas, ni se conoce cómo causan las enfermedades. Lo que sí se sabe es que son replicados por los enzimas del huésped.
- Algunas semejanzas entre la secuencia de nucleótidos de los viroides y ciertos secuencias del genoma de las plantas
no traducidas (intrones) sugieren que los viroides evolucionaron a partir de los intrones.
Leomy Anyelina Porro Mercedes Matricula: 2011-0254 Grupo: 05
ResponderEliminarcontinuación:
OTROS BIONES ACELULARES
Los ARNs satélites son pequeñas moléculas de tamaño similar al de los viroides de plantas (330-400 bases), que son empaquetados en cápsidas de determinadas cepas de virus (con cuyos genomios no muestran homologías). Se replican sólo en presencia del virus colaborador específico, modificando (aumentando o disminuyendo) los efectos patógenos de éste.
Los virusoides constituyen un grupo de ARNs satélites no infectivos, presentes en el interior de la cápsida de ciertos virus, con semejanzas estructurales con los viroides, replicándose exclusivamente junto a su virus colaborador.
LOS PRIONES I y II
Son entidades infectivas de un tipo totalmente nuevo y original, descubiertas por Stanley Prusiner en 1981, responsables de ciertas enfermedades degenerativas del sistema nervioso central de mamíferos (por ejemplo, el “scrapie” o prurito de ovejas y cabras), incluyendo los humanos (kuru, síndrome de Gerstmann-Straüssler, enfermedad de Creutzfeldt-Jakob). Se definen como pequeñas partículas proteicas infectivas que resisten la inactivación por agentes que modifican ácidos nucleicos, y que contienen como componente mayoritario (si no único) una isoforma anómala de una proteina celular. Tanto la versión celular normal (PrPC) como la patógena (PrPSc en el caso del “scrapie”) son glicoproteínas codificadas por el mismo gen cromosómico, teniendo la misma secuencia primaria. Se desconoce si las características distintivas de ambas isoformas estriban en diferencias entre los respectivos oligosacáridos que adquieren por procesamiento post-traduccional.
A diferencia de los virus, los priones no contienen ácido nucleico y están codificados por un gen celular. Aunque se multiplican, los priones de nueva síntesis poseen moléculas de PrP que reflejan el gen del hospedador y no necesariamente la secuencia de la molécula del PrP que causó la infección previa. Se desconoce su mecanismo de multiplicación, y para discernir entre las diversas hipótesis propuestas quizá haya que dilucidar la función del producto normal y su posible conversión a la isoforma patógena infectiva.
Recientemente se ha comprobado que, al menos algunas de las enfermedades por priones son simultáneamente infectivas y genéticas, una situación insólita en la Patología humana, habiéndose demostrado una relación entre un alelo dominante del PrP y la enfermedad de Creutzfeldt-Jakob. El gen del prión (Prn-p) está ligado genéticamente a un gen autosómico (Prn-i) que condiciona en parte los largos tiempos de incubación hasta el desarrollo del síndrome.
¡GRACIAS! :D
#1 Mat: 2011-0289 ..
ResponderEliminarLa Bacteriología es la rama de la Biología que estudia la morfología, ecología, genética y bioquímica de las bacterias así como otros muchos aspectos relacionados con ellas. Es de gran importancia para el hombre por sus implicaciones médicas, alimentarias y tecnológicas.
#2 Mat:2011-0289 -Origen de las celulas Eucariotas-
El origen de los eucariontes es un complejo proceso que tiene un origen procariota. Si bien hay varias teorías que explican este proceso, según la mayoría de estudios se produjo por endosimbiosis entre varios organismos procariotas, en donde el ancestro principal protoeucariota es de tipo arqueano y las mitocondrias y cloroplastos son de origen bacteriano.
#3 Mat:2011-0289 -LA CELULA BACTERIANA-
Las bacterias son microorganismos unicelulares que presentan un tamaño de unos pocos micrómetros (entre 0,5 y 5 μm, por lo general) y diversas formas incluyendo esferas (cocos), barras (bacilos) y hélices (espirilos). Las bacterias son procariotas y, por lo tanto, a diferencia de las células eucariotas (de animales, plantas, hongos, etc.), no tienen el núcleo definido ni presentan, en general, orgánulos membranosos internos. Generalmente poseen una pared celular compuesta de peptidoglicano. Muchas bacterias disponen de flagelos o de otros sistemas de desplazamiento y son móviles.
Las bacterias presentan una amplia variedad de tamaños y formas. La mayoría presentan un tamaño diez veces menor que el de las células eucariotas, es decir, entre 0,5 y 5 μm.
#4 Mat:2011-0289 -La pared gram positiva y gram negativa-
La gram positiva mide de unos 15 a 20 nanometros de diametro y la gram negativa mide 2 nanometros de diametro.
La pared gram positiva, poseen una pared celular interna y una pared de peptidoglucano(es un exoesqueleto que da consistencia y forma esencial para replicación y supervivencia de la bacteria), no tiene membrana externa, no tiene espacio periplasmático, la red de mureína está muy desarrollada y llega a tener hasta 40 capas, conservan el complejo yodocolorante.
La pared gram negativa, poseen una pared celular más compleja:
pared celular interna, pared de peptidoglucano, bicapa lipídica externa, tiene membrana externa que forma un saco rígido alrededor de la bacteria, mantiene estructura y es barrera impermeable a macromoléculas, ofrece protección en condiciones adversas, tiene espacio periplasmático, pueden ser anaerobios o aerobios y poseen proteínas con concentraciones elevadas.
#5 Mat:2011-0289 -COLORACION DE GRAM-
ResponderEliminarLa tinción de Gram o coloración de Gram es un tipo de tinción diferencial empleado en Bacteriología para la visualización de bacterias, sobre todo en muestras clínicas. Debe su nombre al bacteriólogo danés Christian Gram, que desarrolló la técnica en 1884.
Se utiliza tanto para poder referirse a la morfología celular bacteriana como para poder realizar una primera aproximación a la diferenciación bacteriana, considerándose Bacteria Gram positiva a las bacterias que se visualizan de color moradas y Bacteria Gram negativa a las que se visualizan de color rosa o rojo o grosella.
Metodologia:
Recoger muestras.
Hacer el extendido en espiral.
Dejar secar a temperatura ambiente o fijarlas utilizando un mechero.
Fijar la muestra con metanol durante un minuto o al calor (flameado 3 veces aprox.)
Agregar azul violeta (cristal violeta o violeta de genciana) y esperar 1 minuto. Todas las células gram positivas se tiñen de color azul-púrpura.
Enjuagar con agua.
Agregar lugol y esperar entre 1 minuto.
Enjuagar con agua.
Agregar acetona y/o alcohol y esperar de 8 a 15 segundos aproximadamente (parte crítica de la coloración).
Enjuagar con agua.
Tinción de contraste agregando safranina o fucsina básica y esperar 45 segundos. Este tinte dejará de color rosado-rojizo las bacterias Gram negativas.
#6 Mat:2011-0289 -Ribosomas bacterianos-
Los ribosomas son las estructuras supramoleculares encargadas de la síntesis de proteínas, en un proceso conocido como traducción. La información necesaria para esa síntesis se encuentra en el ARN mensajero (ARNm), cuya secuencia de nucleótidos determina la secuencia de aminoácidos de la proteína; a su vez, la secuencia del ARNm proviene de la transcripción de un gen del ADN. El ARN de transferencia lleva los aminoácidos a los ribosomas donde se incorporan al polipéptido en crecimiento.
#7 Mat:2011-0289 -FORMAS Y AGRUPACIONES BACTERIANAS-
ResponderEliminarLas bacterias normalmente se multiplican por fisión transversal binaria. En muchas especies, las células hijas resultantes de un evento de división por fisión tienden a dispersarse por separado al medio, debido a la actuación de fuerzas físicas (movimiento browniano, cizallamiento, corrientes de convección, etc).
Los principales tipos de formas bacterianas son:
-Cocos (células más o menos esféricas)
-Bacilos (en forma de bastón, alargados),que a su vez pueden tener varios aspectos:cilíndricos,fusiformes y en forma de maza, etc.
Atendiendo a los tipos de extremos, éstos pueden ser:
-Redondeados (lo más frecuente)
-Cuadrados
-Biselados
-Afilados
-Espirilos: al igual que los bacilos, tienen un eje más largo que otro, pero dicho eje no es recto, sino que sigue una forma de espiral, con una o más de una vuelta de hélice.
*Vibrios: proyectada su imagen sobre el plano tienen forma de coma, pero en el espacio suelen corresponder a una forma espiral con menos de una vuelta de hélice.
Otros tipos de formas son filamentos, ramificados o no,anillos casi cerrados, y formas con prolongaciones (con prostecas).
#8 Mat:2011-0289 -AGRUPACIONES DE COCOS-
Algunos géneros bacterianos se agrupan de una manera característica. Esta agrupación se debe a la tendencia de las células hijas a permanecer parcialmente adheridas después de la división celular.
Los cocos pueden disponerse:
-De a pares y se los llama diplococos
-Si se disponen en cadena se llaman estreptococos
-Cuatro células esféricas conforman una tetrada
-En forma de racimo o irrregular se llaman estafilococos
-En paquetes cúbicos se denominan sarcinas
#9 Mat:2011-0289 -FLAGELOS Y ESPORAS-
ResponderEliminar-Esporas
Espora en biología designa una célula reproductora generalmente haploide y unicelular. La reproducción por esporas permite al mismo tiempo la dispersión y la supervivencia por largo tiempo (dormancia) en condiciones adversas.
La espora produce un nuevo organismo al dividirse por mitosis sin fusión con otra célula, produciendo un gametofito pluricelular. La espora es un elemento importante en los ciclos vitales biológicos de plantas, hongos y algas. El término deriva del griego σπορά (sporá), "semilla". Las esporas se pueden clasificar según su función, estructura, origen del ciclo vital o por su movilidad.
El término también puede referirse a la etapa inactiva de algunas bacterias, lo que se denomina más correctamente endosporas y no son esporas en el sentido considerado aquí. La mayoría de los hongos producen esporas; aquellos que no lo hacen se denominan hongos asporógenos.
-Flagelos
Son filamentos proteicos responsables de la movilidad. Están constituidos por una proteína llamada flagelina. Se introducen en la membrana celular y se proyectan hacia fuera de la célula.
Los flagelos se disponen de distinta manera alrededor del cuerpo bacteriano.
En algunas especies se encuentran en gran número y distribuídos sobre toda la superficie celular, a esta disposición se la llama peritrica. Cuando la bacteria tiene un único flagelo, en uno de los polos se denomina monotrica. Otras presentan un haz de flagelos en uno de los polos, se las llama lofotrica, las que presentan haces de flagelos en ambos polos, se las llama anfitricas, y las especies que carecen de flagelos se las denomina atricas.
Los flagelos mueven a la célula mediante rotación y le permiten a la bacteria responder a nutrientes químicos o materiales tóxicos y acercarse o alejarse de ellos por el fenómeno llamado quimiotáxis.
Ciertas bacterias usan otros medios de locomoción, las espiroquetas se mueven pero no poseen flagelos, presentan un filamento axial, que les permite el movimiento. Otras como las Mixobacterias, lo hacen por deslizamiento.
#10 Mat:2011-0289 -GENEROS BACTERIANOS-
ResponderEliminarGéneros bacterianos
I. Aeróbicas y Gram negativas, cocos y bacilos
Familias
1. Pseudomonaceae son bacilos curvos o rectos, mótiles por flagelo polar, catalasa y oxidasa positiva. Contribuyen a degradar substancias quíímicas en el suelo, como por ejemplo pesticidas.
Aquí encontramos el género Pseudomonas que se encuentra ampliamente distribuido en la tierra y en el agua. En este grupo incluimos la especie P. syringae que es un patógeno vegetal y causante de tumores a plantas.
Xanthosomas todas las especies de este género son patógenas vegetales.
2. Rhizobiaceae Esta familia ocasiona hipertrofia en plantas, además de la formación de nódulos en raíces. Algunas especies pueden ocasionar tumores.
Rhizobium forma nódulos en raíces de leguminosas y fija N2 atmosférico.
Agrobacterium es importante en la ingeniería genética. Esta posee un plásmido con una región llamada T DNA que se inserta en el genoma de la planta huésped.
3. Methylococcaceae posee la habilidad para usar gas metano como fuente de carbono y energía bajo condiciones aeróbias o microaerofílicas.
4. Acetobacteraceae esta familia oxida etanol a ácido acético.
Acetobacter y Gluconobacter son saprófitos que se encuentran en medios acídicos enriquecidos con azúcar o alcohol como flores, frutas, cervezas, vino, vinagre y otros. Poseen importancia industrial, ya queAzotobacter produce vinagre y Gluconobacter se utiliza para la manufactura de productos químicos.
5. Legionellaceae
Legionella son oportunistas en humanos, pueden causar la muerte. Se transmiten por vía aérea
6. Neisseriaceae
Neisseria son parásitos que habitan en las membranas mucosas de humanos y animales. N. gonorrhoeae y N. meningitidis son altamente patógenos, y este último causa meningitis cerebroespinal.
Acetobacter son saprófitos que se encuentran en tierra, agua y basura. Son patógenos oportunistas que ocasionan una variedad de infecciones particularmente en pacientes hospitalizados.
II. Anaeróbico facultativo, bacilos y gram negativo
#12 Mat:2011-0289 -PRECURSORES BIOSINTETICOS BACTERIANOS-
ResponderEliminar-La glucosa-6-fosfato (glucosa 6P) interviene en las principales rutas del metabolismo de los glúcidos.
-El ácido oxalacético o su forma ionizada, el oxalacetato, es un importante metabolito intermediario de varias rutas metabólicas, entre ellas el ciclo de Krebs y la fotosíntesis C-4. Se puede sintetizar por dos reacciones anapleróticas al pasar el piruvato a oxalacetato por carboxilación o de aspartato a oxalacetato por transaminación.
-El ácido fosfoenolpirúvico es un ácido carboxílico. Su anión se denomina fosfoenolpiruvato y posee gran importancia en el metabolismo celulardebido a que posee un enlace fosfato de alta energía implicado en la glucólisis y gluconeogénesis.
-La enzima Alfa-cetoglutarato deshidrogenasa u oxoglutarato deshidrogenasa (succinil transferidora), EC 1.2.4.2, cataliza la siguientereacción utilizando como cofactor la tiamina difosfato.
#13 Mat:2011-0289 -FORMAS L-
-Formas L:
Protoplastos y Esperoplastos
Diferencias con los micoplasmas
Se denominan protoplastos las células bacterianas a las que se ha desprovisto totalmente de pared celular, mientras que esferoplastos son aquellas células bacterianas que poseen restos de pared.En el caso de Gram-positivas, la desorganización total de su pared, por lo que se obtienen protoplastos; en el caso de las Gram-negativas, quedan restos de membrana externa y de peptidoglucano atrapados en ella, por lo que se obtienen esferoplastos.
A diferencia de los micoplasmas son bacterias que carecen de pared celular. Debido a la ausencia de pared celular, los micoplasmas no son sensibles a los antibióticos que bloquean la síntesis de la pared celular, como la penicilina u otros antibióticos betalactámicos.
#14 Mat:2011-0289 -NUTRICION BACTERIANA-
ResponderEliminarNutricion bacteriana (Matricula 2011-0546)
Las bacterias, como el resto de los seres vivos, necesitan una fuente de carbono para poder sobrevivir. El origen de esta fuente de carbono sirve como criterio de clasificación para las bacterias. Además, se necesita una fuente de energía que sirva para poder construir sus propias moléculas; el tipo de fuente de energía utilizada también sirve como criterio de clasificación.
Autótrofas: la fuente de carbono es inorgánica (CO2).
Fotolitotrofas: la energía utilizada es la luz. (Ejemplo: bacterias purpúreas del azufre).
Quimiolitotrofas: la energía utilizada es la liberada en reacciones químicas. (Ejemplo: bacterias incoloras del azufre).
Heterótrofas: la fuente de carbono es inorgánica.
Fotoorganotrofas: la energía utilizada es la luz.
Quimioorganotrofas: la energía utilizada es la liberada en reacciones químicas. A este grupo pertenecen la mayoría de las bacterias.
Bacterias aerobias: son aquellas que necesitan oxígeno para su metabolismo. Realizan la oxidación de la materia orgánica en presencia de oxígeno molecular, es decir, realizan la respiración celular.
Bacterias anaerobias: son aquellas que no utilizan oxígeno molecular en su actividad biológica. La obtención de energía la realizan mediante catabolismo fermentativo. Se pueden distinguir dos grupos dentro de ellas:
Bacterias anaerobias facultativas: pueden vivir en ambientes con oxígeno o sin él.
Bacterias anaerobias estrictas: sólo pueden sobrevivir en ambientes carentes de oxígeno. Como ejemplo, Clostridium, causante del tétanos.
#17 Mat:2011-0289 -Mecanismo de resistencia bacteriana-
ResponderEliminarLa mayoría de bacterias que hacen resistencia a algún antibiótico tienen la capacidad de eludir o evitar la acción del medicamento, gracias a 5 mecanismos:
-Producción de enzimas que destruyen fármaco activo: Este mecanismo es utilizado principalmente por los Staphylococos resistentes a la penicilina G, gracias a la producción de B lactamasas que destruyen el fármaco.
-Cambio de permeabilidad al fármaco: Algunas bacterias como los estreptococos poseen una barrera natural de permeabilidad a los aminoglucósidos, que puede ser superada al administrar simultáneamente un fármaco activo contra la pared celular como por ejemplo la penicilina.
-Alteran estructuralmente el "blanco del fármaco" , sucede principalmente con antibióticos como penicilinas y cefalosporinas, en las que la resistencia es ocasionada por alteración o perdida de la función de las PBP (Proteínas de unión a penicilinas).
-Desarrollo de via metabólica diferente que pasa por alto la reacción inhibida por el fármaco: Hay bacterias que son resistentes a las sulfonamidas y no requieren PABA extracelular y, pueden utilizar Ácido fólico preformado
-Desarrollo de enzima diferente que ejecuta función metabólica pero es menos afectada por el fármaco. Un ejemplo claro de esto se presenta en las bacterias resistentes al trimetropim, la ácido dihidrofolico reductasa se inhibe con mucha menor eficiencia que en bacterias susceptibles al trimetropim.
#1 Mat:2011-0289 -Mecanismo de Acción de Los Antibióticos-
ResponderEliminarDebido a que los antibióticos tienen efectos sobre una diversidad de bacterias, sus mecanismos de acción difieren basados en las características vitales de cada organismo y que, por lo general, son objetivos que no existen en las células de mamíferos.
Los agentes antimicrobianos actúan por una serie de mecanismos, muy diferentes entre ellos y cuyos blancos se encuentran en diferentes regiones de la célula atacada. Las diversas regiones de ataque antibacteriano en general son consideradas:
* Pared bacteriana
* Membrana bacteriana
* Síntesis de proteínas
* Síntesis de ácidos nucleicos
La acción del agente antibacteriano es lograda mediante los siguientes mecanismos de acción:
* inhibición de la síntesis de la pared celular
* inhibición de la síntesis de proteínas
* inhibición del metabolismo bacteriano
* inhibición de la actividad o síntesis del ácido nucleico
* alteraciones en la permeabilidad de la membrana celular
Con cualquiera de estas acciones o con una combinación de ellas, el germen es incapaz de sobrevivir.
#2 Mat:2011-0289 -PENICILINAS-
ResponderEliminarLas penicilinas son antibióticos del grupo de los betalactámicos empleados profusamente en el tratamiento de infecciones provocadas por bacterias sensibles. La mayoría de las penicilinas son derivados del ácido 6-aminopenicilánico, difiriendo entre sí según la sustitución en la cadena lateral de su grupo amino. La penicilina G o bencipenicilina fue el primer antibiótico empleado ampliamente en medicina; su descubrimiento ha sido atribuido a Alexander Fleming en 1928, quien junto con los científicos Ernst Boris Chain y Howard Walter Florey —que crearon un método para producir en masa el fármaco— obtuvo el Premio Nobel en Fisiología o Medicina en 1945.
Existe una gran diversidad de penicilinas. Algunas especies de hongos del género Penicillium sintetizan de forma natural penicilinas, como el primer tipo aislado, la penicilina G. No obstante, debido a la aparición de resistencias, se han desarrollado otras familias siguiendo básicamente dos estrategias: la adición de precursores para la cadena lateral en el medio de cultivo del hongo productor, lo que se traduce en la producción de penicilinas biosintéticas; y la modificación química de la penicilina obtenida por la fermentación biotecnológica, lo que da lugar a las penicilinas semisintéticas.
La penicilina natural o penicilina G es cristalina, totalmente soluble en agua, soluciones salinas y dextrosas isotónicas. El radical R, es el responsable de la sensibilidad a la hidrólisis por parte de las β-lactamasas, del enlace a proteínas transportadoras y del vínculo con las proteínas bacterianas PBP que transportan a la penicilina dentro de la célula.37 Además, se le asocia a la penicilina un dipéptido cisteína-valina, haciendo que la penicilina tenga la ideal afinidad por la enzima bacteriana transpeptidasa, la cual no se encuentra en el cuerpo humano y que permite la síntesis del peptidoglucano.
#3 Mat:2011-0289
ResponderEliminarLas cefalosporinas, son una clase de los antibióticos beta-lactámicos. Junto con las cefamicinas pertenecen a un subgrupo llamado los cefamos. Las cefalosporinas son similares a las penicilinas, pero más estables ante muchas β-lactamasas bacterianas y, por lo tanto, tienen un espectro de actividad más amplio.
as cefalosporinas son agrupadas en grupos llamados "generaciones" por sus características antimicrobianas. Las primeras cefalosporinas fueron agrupadas en la "primera generación" mientras que más adelante, cefalosporinas de espectro extendido fueron clasificadas como cefalosporinas de segunda generación. Cada nueva generación de cefalosporinas tiene más potencia frente a bacterias gram-negativas, características antimicrobianas perceptiblemente mayores que la generación precedente; actualmente se diferencian cuatro generaciones de cefalosporinas. Cabe destacar que las cefalosporinas de primera generación tienen mayor espectro de acción ante estafilococo y estreptococo que las generaciones más recientes.
Se clafisican:
Cefalosporinas de primera generación
Las cefalosporinas de primera generación tienen actividad predominante contra cocos grampositivos —Streptococcus y Staphylococcus—. Las cefalosporinas usuales no son activas contra cepas de Staphylococcus aureus resistentes a meticilina (SARM).
Cefalosporinas de segunda generación
Desde esta generación se amplia el espectro incluyendo anaerobios y microorganismos gram negativos. Las bacterias del género Klebsiella suelen ser sensibles
Cefalosporinas de tercera generación
Cefdinir
Cefpodoxima
Cefditoren pivoxilo
Cefixima
Cefalosporinas de cuarta generación
Cefalosporinas de la cuarta generación tienen un mayor espectro de la actividad contra organismos gram-positivos que las cefalosporinas de la tercera generación. También tienen una mayor resistencia a beta-lactamasas que las cefalosporinas de la tercera generación.
Cefetecol (Cefcatacol)
Cefquinome (Cephaguard) (*uso veterinario*)
Flomoxef (Flumarin)
Cefalosporinas de quinta generación
Ceftobiprol : mayor actividad contra Staphylococccus aureus resistentes a meticilina, Streptococcus pneumonie resistente a penicilinas, Pseudomona aeruginosa y enterococos.
Ceftarolina : activa tanto frente a Staphylococcus aureus como estafilococos coagulasa negativos sensible y resistente a meticilina (MRSA) así como frente a las recientes cepas resistentes a vancomicina (VRSA) y resistentes a daptomicina
#4 Mat:2011-0289
ResponderEliminarTetraciclinas, Amino glúcidos, Imidazoles, Macrolidos, Lincosamidas y Sulfamidas
La tetraciclina: es usada para tratar las infecciones provocadas por bacterias, incluyendo la neumonía y otras infecciones en las vías respiratorias; el acné; infecciones en la piel, los genitales y el sistema urinario; y la infección que causa úlceras estomacales (Helicobacter pylori). Este medicamento es también usado como una alternativa de otros medicamento para el tratamiento de la enfermedad de Lyme y para el tratamiento y prevención del carbunco (después de una exposición por inhalación). La tetraciclina pertenece a una clase de medicamentos llamados antibióticos de tetraciclina. Funciona al prevenir el crecimiento y diseminación de las bacterias. Los antibióticos no tienen ningún efecto en los resfríos, la gripe u otras infecciones víricas.
Los aminoglucósidos o aminósidos: son un grupo de antibióticos bactericidas que detienen el crecimiento bacteriano actuando sobre sus ribosomas y provocando la producción de proteínasanómalas. Actúan a nivel de ribosomas en la subunidad 30S bacteriana, y por ende, a nivel de síntesis de proteínas, creando porosidades en la membrana externa de la pared celular bacteriana. Tienen actividad especialmente en contra de bacterias Gram negativas y aeróbicas y actúan sinergísticamente en contra de organismos Gram positivos.
El imidazol: es un intermediario de la biosíntesis de la histidina que se forma desde el imidazol glicerol fosfato con la pérdida de agua. De estructura cristalina, su fórmula molecular es C3H4N2 y su masa molecular es 68.077 g/mol.
Pertenece a una familia de compuestos químicos aromáticos, caracterizada por disponer de un anillo pentagonal nitrogenado. Tiene propiedades anfotéricas, cualidad esta que mantendrán algunos de sus derivados.
Los macrólidos: son antibióticos naturales, semisintéticos y sintéticos que ocupan un lugar destacado en el tratamiento de infecciones causadas por bacterias intracelulares. Integran este grupo: eritromicina, claritromicina, azitromicina, espiramicina y roxitromicina.
Lincosamidas: Son antibióticos bacteriostáticos o bactericidas en dependencia de su concentración, el uso de este medicamento por v.i ha obtenido magníficos resultados en el tratamiento para bacterias anaerobias.
Una sulfamida : es una sustancia química sintética derivada de la sulfonamida, bacteriostática y de amplio espectro. Las sulfamidas se emplean como antibióticos,antiparasitarios y coccidiostáticos en el tratamiento de enfermedades infecciosas. Las sulfas son usadas tanto en medicina humana, como el sulfametoxazol, la sulfadiazina y la sulfabenzamida, así como veterinaria, como el sulfanitrato, el sulfapirazol y la sulfaquinoxalina. Las sulfonamidas fueron las primeras drogas eficaces empleadas para el tratamiento sistémico de infecciones bacterianas en el ser humano.
#5 Mat:2011-0289
ResponderEliminarTetraciclinas
Las tetraciclinas constituyen un grupo de antibióticos, unos naturales y otros obtenidos por semisíntesis, que abarcan un amplio espectro en su actividad antimicrobiana. Químicamente son derivados de la naftacenocarboxamida policíclica, núcleo tetracíclico, de donde deriva el nombre del grupo.
Las tetraciclinas naturales se extraen de las bacterias del género Actinomyces. De Streptomyces aurofaciens se extraen la clortetraciclina y la demetilclortetraciclina, de Streptomyces rimosus se extrae la oxitetraciclina, y la tetraciclina, representante genérico del grupo, se puede extraer del Streptomyces viridifaciens, aunque también se puede obtener de forma semisintética. Una característica común al grupo es su carácter anfotérico, que le permite formar sales tanto con ácidos como con bases, utilizándose usualmente los clorhidratos solubles.
Actúan fundamentalmente como bacteriostáticos a las dosis habituales, aunque resultan bactericidas a altas dosis, generalmente tóxicas. Actúan por varios mecanismos:
Desacoplan la fosforilación oxidativa de las bacterias.
Provocan una inhibición de la síntesis protéica en el ribosoma de la bacteria. Actúan inhibiendo la síntesis proteica al unirse a la subunidad 30 S del ribosoma y no permitir la unión del ácido ribonucléico de Transferencia (tRNA) a este, ni el transporte de aminoácidos hasta la subunidad 50 S.1
Existe también evidencia preliminar que sugiere que las tetraciclinas alteran la membrana citoplasmática de organismos susceptibles, permitiendo la salida de componentes intracelulares.5
Las resistencias bacterianas a las tetraciclinas son de aparición lenta, aunque mucho más rápida si se utiliza por vía tópica. El mecanismo bacteriano implicado puede ser mediante plásmido, lo que explica la reticencia a usar las tetracilinas en el ámbito hospitalario, para evitar la aparición de resistencias simultáneas a varios antibióticos. Existen resitencias cruzadas entre los miembros del grupo.
-Se clasifican en :
Demeclociclina
Doxiciclina
Clortetraciclina:
Metaciclina
Oxitetraciclina
Minociclina
Rolitetraciclina
A. Viroides
ResponderEliminar- Pequeñas moléculas de ARN circular de cadena sencilla formadas por unos pocos cientos de nucleótidos. Carecen de
cáps ida y su tamaño es una milésima parte del de los virus más pequeños.
- Aparecen con mayor abundancia en el núcleo de las células infectadas.
- Sólo han sido detectados en plantas, en las que producen una gran variedad de enfermedades y pueden ser transmitidos
tanto horizontal como verticalmente.
- No existen ninguna evidencia de que los viroides son traducidos a proteínas, ni se conoce cómo causan las enfermedades.
Lo que sí se sabe es que son replicados por los enzimas del huésped.
- Algunas semejanzas entre la secuencia de nucleótidos de los viroides y ciertos secuencias del genoma de las plantas
no traducidas (intrones) sugieren que los viroides evolucionaron a partir de los intrones.
C. Priones
- Las encefalopatías espongiformes transmisibles (TSEs) son enfermedades cerebrales degenerativas que ocurren en
muchos mamíferos, incluido el hombre. En estas enfermedades, se desarrollan progresivamente cavidades que hacen
que adquiera el aspecto de una esponja. El scrapie es una TSE que afecta a ovejas y cabras conocida desde hace
250 años.
- En los años 80 aparecieron numerosos casos de vacas afectadas por una TSE en Gran Bretaña que pudieron relacionarse
con la ingestión de piensos obtenidos a partir de ovejas afectadas por el scrapie. En los 90 apareció una variedad
de la enfermedad que afectaba a algunas personas que habían consumido carne de vacas con TSE y que los
medios de comunicación bautizaron con el nombre de “mal de las vacas locas”.
- El kuru es otra TSE que afectaba a los nativos de una tribu de Nueva Guinea que practicaban canibalismo ritual.
· Los priones son partículas infecciosas proteicas que provocan las encefalopatías espongiformes transmisibles.
· Poseen la misma secuencia aminoacídica de una proteína normal, pero presentan una estructura diferente.
· Inducen, por un mecanismo hasta ahora desconocido, la transformación de proteínas normales en anómalas.
· Contradicen el dogma central de la biología molecular, según el cual el flujo de información es en todos los seres vivos:
ADN à ARNm à Secuencia Aminoácidos à Estructura Tridimensional Proteínas. La teoría de los priones propuesta
por Prusiner supone la existencia de dos plegamientos para una única secuencia de amino ácidos. Además, el
replegamiento de la PrP normal por acción de la PrP patológica, implica un flujo de información de una proteína a otra
a nivel de estructura terciaria.
DAYBELYS SANTANA 86166
ResponderEliminar#6 RIBOSOMAS BACTERIANOS
Los ribosomas son complejos macromoleculares de proteínas y ácido ribonucleico (ARN) que se encuentran en el citoplasma, en las mitocondrias, en el retículo endoplasmatico y en los cloroplastos. Son un complejo molecular encargado de sintetizar proteínas a partir de la información genética que les llega del ADN transcrita en forma de ARN mensajero (ARNm). Sólo son visibles al microscopio electrónico, debido a su reducido tamaño (29 nm en células procariotas y 32 nm en eucariotas). Bajo el microscopio electrónico se observan como estructuras redondeadas, densas a los electrones. Bajo el microscopio óptico se observa que son los responsables de la basofilia que presentan algunas células. Están en todas las células (excepto en los espermatozoides). Los ribosomas no se definen como orgánulos, ya que no existen endomembranas en su estructura.[cita requerida]
En células eucariotas, los ribosomas se elaboran en el núcleo pero desempeñan su función de síntesis en el citosol. Están formados por ARN ribosómico (ARNr) y por proteínas. Estructuralmente, tienen dos subunidades. En las células, estas macromoléculas aparecen en diferentes estados de disociación. Cuando están completas, pueden estar aisladas o formando grupos (polisomas). Las proteínas sintetizadas por los ribosomas actúan principalmente en el citosol; también pueden aparecer asociados al retículo endoplasmático rugoso o a la membrana nuclear, y las proteínas que sintetizan son sobre todo para la exportación.
Tanto el ARNr como las subunidades de los ribosomas se suelen nombrar por su coeficiente de sedimentación en unidades Svedberg. En las células eucariotas, los ribosomas del citoplasma alcanzan 80 S. En plastos de eucariotas, así como en procariotas, son 70 S. Los ribosomas mitocondriales son de tamaño variado, entre 55 y 80 S.1
DAYBELYS SANTANA 86166
ResponderEliminar#DIAPOSITIVA 1 BACTERIOLOGIA
La Bacteriología es la rama de la Biología que estudia la morfología, ecología, genética y bioquímica de las bacterias así como otros muchos aspectos relacionados con ellas. Es de gran importancia para el hombre por sus implicaciones médicas, alimentarias y tecnológicas
La bacteriología se clasifica en: • Bacteriología General • Bacteriología Agrícola • Bacteriología Alimenticia • Bacteriología Farmacéutica • Bacteriología Médica
Hay diversas clasificaciones para un mejor estudio de las bacterias: • Morfología • Organización celular • Presencia de núcleo • Tipo de nutrición • Tipo de reproducción • Tinción de Gram
La bacteriología se clasifica en: • Bacteriología General • Bacteriología Agrícola • Bacteriología Alimenticia • Bacteriología Farmacéutica • Bacteriología Médica
Hay diversas clasificaciones para un mejor estudio de las bacterias: • Morfología • Organización celular • Presencia de núcleo • Tipo de nutrición • Tipo de reproducción • Tinción de Gram
Hay diversas clasificaciones para un mejor estudio de las bacterias: • Morfología • Organización celular • Presencia de núcleo • Tipo de nutrición • Tipo de reproducción • Tinción de Gram
Cynthia Pamela de la Rosa --------------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3
ResponderEliminarDIAPOSITIVA: #1
QUE ES LA VIROLOGIA?
A traves de mis investigaciones pude encontrar la importancvia de la virologia, que es?, entre otros temas sobre la misma, a continuacion su definicion.
Virología es el estudio de virus y virus como agentes: clasificación y evolución, su estructura, sus formas de infectar y explotar las células para la reproducción del virus, las enfermedades que causan, las técnicas para aislar y la cultura de ellos y su uso en la investigación y terapia.
Virología es considerada una parte de la microbiología o de patología.
Una importante rama de la Virología es la clasificación de virus. Virus pueden clasificarse según la célula huésped que infectan: virus animales, virus de plantas, hongos virus y bacteriófagos (virus que infectan bacterias, que son los virus más complejos).
Otra clasificación utiliza la forma geométrica de su cápside (a menudo una hélice o un icosaedro) o la estructura del virus (por ejemplo, la presencia o la ausencia de un envolvente de lípidos). Gama de virus en tamaño de unos 30 nm a aproximadamente 450 nm, lo que significa que la mayoría de ellos no puede verse con microscopios de luces.
Cynthia Pamela de la Rosa ----------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3
ResponderEliminarDIAPOSITIVA: #2
ESTRUCTURAS GENERALES DE LOS VIRUS
Sabemos que un virus está compuesto de una molécula de ácido nucleico y una envoltura proteínica. Ésta es la estructura básica de un virus, aunque algunos de ellos pueden añadir a esto la presencia de alguna enzima, bien junto al ácido nucleico, como la transcriptasa inversa de los retrovirus, bien en la envoltura, para facilitar la apertura de una brecha en la membrana de la célula hospedadora.
A la unidad formada por el ácido nucleico y la envoltura proteínica se le denomina también virión.
El ácido nucleico es solamente de un tipo, ADN o ARN, nunca los dos.
Atendiendo al tipo de ácido nucleico se distinguen cuatro clases de virus:
- ADN de cadena doble
- ADN de cadena sencilla
- ARN de cadena doble
- ARN de cadena sencilla
La envoltura proteínica recibe el nombre de cápsida. Está formada por unas subunidades idénticas denominadas capsómeros. Los capsómeros son proteínas globulares que en ocasiones tienen una parte glicídica unida. Se ensamblan entre sí dando a la cubierta una forma geométrica.
DIOS LO BENDIGA :D
Cynthia Pamela de la Rosa ----------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3
ResponderEliminarDIAPOSITIVA: #3
SIMETRIA VIRALES
Atendiendo la forma de la cápsida, se pueden distinguir los siguientes tipos de virus:
- Cilíndricos o helicoidales: los capsómeros, que son de un solo tipo, se ajustan entorno una hélice simple de ácido nucleico. Un ejemplo lo constituye el virus del mosaico del tabaco.
- Icosaédricos: los capsómeros, que suelen ser de varios tipos, se ajustan formando un icosaedro regular (es decir, 20 caras triangulares y 12 vértices), y dejando un hueco central donde se sitúa el ácido nucleico fuertemente apelotonado. Algunos forman poliedros con más caras que el icosaedro, y algunos presentan fibras proteicas que sobresalen de la cápsida.
Un ejemplo lo constituyen los adenovirus, entre los que se encuentran los virus de los resfriados y faringitis.
- Complejos: con pequeñas variantes, responden a la siguiente estructura general:
- Una cabeza de estructura icosaédrica que alberga el ácido nucleico.
- Una cola de estructura helicoidal que constituye un cilindro hueco.
- Un collar de capsómeros entre la cabeza y la cola.
- Una placa basal, al final de la cola, con unos puntos de anclaje que sirven para fijar el virus a la membrana celular. De la placa salen también unas fibras proteicas que ayudan a la fijación del virus sobre la célula hospedadora.
Como ejemplo de este tipo de virus se encuentran la mayor parte de los virus bacteriófagos (que infectan las bacterias).
Cynthia Pamela de la Rosa ----------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3
ResponderEliminarDIAPOSITIVA: #4
Tamaño de los virus
Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus.
Su pequeño tamaño explica lo tardío del descubrimiento de estos agentes. La primera referencia sobre la existencia de los virus se debe al botánico ruso Dimitri Ivanovski en 1892. Este investigador buscaba el agente causante de la enfermedad denominada mosaico del tabaco, y llegó a la conclusión de que debía tratarse de una toxina o de un organismo más pequeño que las bacterias, pues el agente atravesaba los filtros que retenían las bacterias. Denominó a estos agentes patógenos virus filtrables.
Cynthia Pamela de la Rosa ----------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3
ResponderEliminarDIAPOSITIVA: #5
FAMILIA DE LOS VIRUS y SUS ESPECIES IMPORTANTES
~Adenoviridae....dsDNA....adenovirus..no-encapsulado..
~Picornaviridae... .virus ssRNA sentido-positivo..... coxsackievirus, virus de hepatitis A, poliovirus no-encapsulado
~Herpesviridae....dsDNA.....Virus de Epstein-Barr, herpes simple, tipo 1, herpes simplex, tipo 2, citomegalovirus humano, herpesvirus humano, tipo 8, Virus varicela-zoster.... encapsulado
~Hepadnaviridae dsDNA y ssDNA...... virus de hepatitis B encapsulado
~Flaviviridae...... virus ssRNA sentido-positivo..... virus de hepatitis encapsulado
~Retroviridae..... virus ssRNA sentido-positivo..... Virus de inmunodeficiencia humana (VIH) encapsulado
~Orthomyxoviridae...... virus ssRNA sentido-negativo..... virus de influenza encapsulado
~Paramyxoviridae..... virus ssRNA sentido-negativo..... virus del sarampión, virus de parotiditis, virus de parainfluenza, virus sincitial respiratorio encapsulado
~Papovaviridae ssDNA..... papilomavirus..... no-encapsulado
~Rhabdoviridae virus ssRNA sentido-negativo virus de la rabia encapsulado
~Togaviridae virus ssRNA sentido-positivo virus de Rubéola encapsulado
BENDICIONES MAESTRO! :D
Cynthia Pamela de la Rosa ----------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3
ResponderEliminarDIAPOSITIVA: #6
DIAGNOSTICO DE LA VIROSIS
* METODOS DIRECTOS
Son aquellos que detectan:
1. El virus como agente infeccioso (aislamiento viral).
2. La presencia de antígenos virales (técnicas inmunológicas): Inmunofluorescencia (IF), Enzimoinmunoanálisis (EIA), Test de Aglutinación.
3.La presencia de ácidos nucleicos virales (PCR, etc.).
4.El virus como partícula viral (microscopía electrónica).
METODOS INDIRECTOS
Son aquellos que reconocen la respuesta inmune (humoral o celular) por parte del huésped: . Detección de anticuerpos específicos antivirales por técnicas inmunológicas (EIA, IFI, WB, etc.). Producción de anticuerpos in vitro.
En el curso de una infección varían las poblaciones de anticuerpos frente al agente infectante. En una primera fase la clase predominante suele ser IgM, mientras que con el transcurso del tiempo las IgM disminuyen hasta desaparecer o quedar a baja concentración residual y, en cambio, aumentan las IgG. La búsqueda de anticuerpos clase IgM es de utilidad para hacer diagnóstico de infección reciente en una sola muestra de suero extraída en el período agudo de la enfermedad. Este método se emplea para el diagnóstico de enfermedades como: Rubéola, Citomegalovirus, Hepatitis a virus A, etc.
Cynthia Pamela de la Rosa ----------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3
ResponderEliminarDIAPOSITIVA: #7
ETAPAS DE REPLICACION DEL VIRUS
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante.
Fijación, penetración y pérdida de la cubierta.
Cynthia Pamela de la Rosa ----------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3
ResponderEliminarDIAPOSITIVA: #8
Penetracion de los Bacteriofagos
Los virus son moléculas de DNA o RNA rodeadas por una envoltura proteica que necesitan células viables para poder replicarse. Los virus utilizan la maquinaria metabólica de las células para sintetizar su material genético y proteínas de la envoltura. Existen distintos tipos de virus que pueden infectar células procariontes o células eucariontes. Los bacteriófagos o fagos son virus que se reproducen en células procariontes.
El genoma de los fagos puede ser RNA simple cadena (MS2, Qß), RNA doble cadena (phi 6), DNA simple cadena (phi X174, fd, M13) o DNA doble cadena (T3, T7, lambda , T5, Mu, T2, T4). Estos ácidos nucleicos pueden contener bases inusuales que son sintetizadas por proteínas del fago. En los T-pares el genoma no contiene citosina sino 5'- hidroximetilcitosina, mientras que en otros tipos de fago alguna de las bases esta parcialmente sustituida.
Cynthia Pamela de la Rosa ----------2011-0385 ( MICROBIOLOGIA I ) GRUPO: #3
ResponderEliminarDIAPOSITIVA: #9
Maduracion y salida de virus envueltos y desnudos.
Los virus cubiertos maduran por un proceso de gemación. Se insertan glucoproteínas de cubierta específica del virus en las membranas celulares y, a continuación, las nucleocápsides hacen gemación a través de la membrana a nivel de estos sitios modificados y, al hacerlo así, adquieren su cubierta. La gemación ocurre a menudo a nivel de la membrana plasmática y puede abarcar a otras membranas de la célula. Los virus cubiertos no son infecciosos hasta que han adquirido sus cubiertas. Por lo tanto, no se acumulan de manera típica viriones descendientes infecciosos en el interior de la célula infectada.
La maduración viral es a veces un proceso ineficaz. Pueden acumularse cantidades excesivas de componentes virales y participar en la formación de cuerpos de inclusión en la célula (ver antes). Como resultado de los profundos efectos dañinos de la replicación viral, se desarrollan por último efectos citopáticos celulares y la célula muere. Sin embargo, hay casos en que el virus no lesiona a la célula y se producen infecciones persistentes y prolongadas.os genomas virales recién sintetizados y los polipéptidos de la cápside se ensamblan para formar los virus descendientes. Las cápsides icosaédricas pueden condensarse en ausencia de ácido nucleico, en tanto que las nucleocápsides de los virus de simetría helicoidal no pueden formarse sin ARN viral.
No existen mecanismos especiales para la liberación de virus carentes de cubierta; las células infectadas acaban por experimentar lisis y liberar las partículas virales.
Este comentario ha sido eliminado por el autor.
ResponderEliminarESTEFANIA ROBLES LAJARA 2012-0358
ResponderEliminarMARTES DE 3-5
Virología esta es una parte de la microbiología y la patología que estudia los virus normal y también como agentes, su clasificación y evolución de los mismos, estructuras sus formas de infectar y explotar las células para la reproducción del virus, las enfermedades que causan, las técnicas para aislar y la cultura de ellos y su uso en la investigación y terapia
Virus pueden clasificarse según la célula huésped que infectan: virus animales, virus de plantas, hongos virus y bacteriófagos (virus que infectan bacterias, que son los virus más complejos).
Otra clasificación utiliza la forma geométrica de su cápside (a menudo una hélice o un icosaedro) o la estructura del virus (por ejemplo, la presencia o la ausencia de un envolvente de lípidos). Gama de virus en tamaño de unos 30 nm a aproximadamente 450 nm, lo que significa que la mayoría de ellos no puede verse con microscopios de luces.
La forma y la estructura del virus ha sido estudiado por cristalografía de rayos X, microscopía electrónica y RMN.
El sistema de clasificación más útil y más ampliamente utilizado distingue virus segun el tipo de ácido nucleico que utilizan como material genético y el método de replicación viral que emplean coaxial células huésped en la producción de más virus:
• Virus de ADN (divididas en virus de ADN bicatenario y los virus de ADN monocatenario mucho menos común),
• Virus ARN (dividieron en virus de ARN monocatenario de sentido positivo, virus de ARN monocatenario de sentido negativo y los virus de ARN bicatenario mucho menos común),
• invertir transcribir virus (virus de ADN de transcripción inversa de bicatenario y monocatenario transcripción inversa RNA virus incluyendo retrovirus).
También estudian '' partículas subvirales '', incluso más pequeñas que los virus infecciosas entidades: viroides (desnudas circulares moléculas de ARN infectando plantas), satélites (moléculas de ácido nucleico con o sin una cápside que requieren un virus auxiliar para la infección y reproducción) y priones (proteínas que pueden existir en una conformación patológica que induce a otras moléculas del prión para asumir esa misma conformación).
Los taxones en Virología no son necesariamente monofiléticos. De hecho, las relaciones evolutivas de los distintos grupos de virus no están claras, y existen tres hipótesis sobre su origen:
1. Virus surgieron de materia no viviente, por separado y en paralelo a otras formas de vida, posiblemente en forma de autorreplicación RNA ribozimas similares a viroides.
2. Virus surgieron de la anteriores, más competentes celulares formas de vida que se convirtieron en parásitos a las células del huésped y posteriormente perdió la mayoría de sus funciones; ejemplos de tales diminutos parásitos procariotas son Mycoplasma y Nanoarchaeum.
3. Virus surgieron como partes del genoma de las células, probablemente transposones o plásmidos, que adquirieron la capacidad para "romper libre" desde el host de celda y infectan otras células.
Por supuesto es posible que diferentes alternativas se aplican a grupos de virus diferente.
De particular interés aquí es mimivirus, un virus gigante que infecta a las amebas y lleva gran parte de la maquinaria molecular tradicionalmente asociada con las bacterias. ¿Es una versión simplificada de una Célula procariota parasitaria o se originó como un simple virus que adquirido genes de su host?
La evolución del virus, que a menudo se produce en concierto con la evolución de sus anfitriones, se estudia en el campo de la evolución viral.
Mientras que el virus se reproduce y evolucionan, ellos no participan en el metabolismo y dependen de una célula huésped para la reproducción. La cuestión debatida a menudo de si están vivos o no es una cuestión de definición que no afectan la realidad biológica de los virus.
continuacion
ResponderEliminarestefania robles 2012-0358
Los viroides al igual que los virus, tienen un ciclo extracelular que se caracteriza por la inactividad metabólica y unciclo intracelular en el que causan infección al huésped susceptible, pero que a diferencia de los virus, los viroides no poseen proteínas ni lípidos y están constituidos por una cadena cíclica corta de ARN, circular o con forma de varilla, (que no codifica proteínas)
Los viroides son los agentes infecciosos de menor complejidad genética y estructural conocidos y representan una forma extrema de parasitismo. Están constituidos exclusivamente por moléculas de ARN de cadena simple, cerradas covalentemente o con forma de bastón de bajo peso molecular (246 a 1994 nucleótidos). Carecen de actividad de ARN mensajero y se replican de forma autónoma, utilizando el sistema de transcripción de la célula suceptible.2 Los viroides se encuentran, casi exclusivamente, en el núcleo de las células infectadas; se desconoce el modo en que se replican pero se sabe que el ARN que los constituyen no funciona como ARN mensajero y tampoco se traduce a enzimas que participen en su propia replicación. Los viroides no codifican para ninguna proteína y llevan a cabo su replicación en la célula hospedadora a través de la RNA polimerasa II codificada por la célula hospedadora aunque también se ha propuesto la posibilidad de que lo haga gracias a una RNA polimerasa dependiente de RNA presente en algunas células eucariotas (Cann, 2001). Dada su localización, se presume que causan la enfermedad interfiriendo con la regulación génica de la célula hospedadora en la etapa de corrección del ARN mensajero celular: particularmente en la eliminación de intrones y de empalmes de exones, presentando secuencias muy similares a los intrones que normalmente son cortados. Sus ARN se replican varias veces.
Un prion1 es un tipo de proteína patógena (es decir, una molécula compleja que induce enfermedades), más específicamente una psialoproteína patógena, la cual tiene alterada su estructura secundaria, teniendo un incorrecto plegamiento de su estructura terciaria.2 A diferencia del resto de los agentes infecciosos (virus, bacterias, hongos etc...), que contienen ácidos nucleicos (ya sea ADN, el ARN, o ambos), un prion sólo esta compuesto por aminoácidos y no presenta material genético. A pesar de que actualmente hay muy poca información sobre la función que tienen las proteínas Prpc (estado no patógeno), algunos experimentos han demostrado que tienen un papel activo en el correcto desarrollo neuronal, que es una proteína capaz de unir específicamente Cu2+ (procesos de oxidoreducción), y también se han relacionado los priones con proteínas de transducción de señales, la adhesión celular y la regulación y distribución de los receptores de acetilcolina.6
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarVIROLOGÍA
La virología (rama de la microbiologia) es el estudio de los virus: su estructura, clasificación y evolución, su manera de infectar y aprovecharse de las células huésped para la reproducción del virus, su interacción con los organismos huéspedes, su inmunidad, la enfermedad que causan, las técnicas para su aislamiento, cultivo y su uso en investigación y terapia. La virología es considerada un sub-campo de la microbiología y la medicina. La virologia también es una rama taxonómica de la biología para estudiar los virus y todos sus componentes.
Su investigación incluye:
• La replicación viral
• Los patógenos virales.
• La inmunología viral.
• Las vacunas virales.
• Los métodos de diagnóstico.
• La quimioterapia antiviral.
• Las medidas de control de una infección.
• Los diferentes signos que manifiestan la presencia de virus.
El Ciclo De Replicación De Los Virus consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden sobrevivir por si mismos en el exterior (véase Virus). Para reproducirse se introducirán en células donde controlarán sus mecanismos reproductivos.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarOrigen de Los Eucariotas
El origen de los eucariotas se encuentra en sucesivos procesos simbiogenéticos (procesos simbióticos que culminan en la unión de sus simbiontes, estableciéndose una nueva individualidad de los integrantes) entre diferentes bacterias. A principios del siglo XX, en 1909, el ruso Kostantin S. Mereschovky presentó la hipótesis según la cual el origen de los cloroplastos tendría su origen en procesos simbióticos.3 A parecidas conclusiones llegaron Kozo-Polyansky y Andrey Faminstyn (también de la escuela rusa) que consideraban la simbiogénesis “crucial para la generación de novedad biológica".4 En Francia, el biólogo Paul Portier, en 1918, y Ivan Wallin en Estados Unidos en 1927, llegaron a las mismas conclusiones. Trabajos que o bien pasaron inadvertidos (como los de la escuela rusa) o no fueron tenidos en cuenta (en el caso de Portier y Wallis) costando el prestigio profesional a sus proponentes. Lynn Margulis rescata estos trabajos y en 1967 en el artículo On origen of mitosing cells presenta la que llegaría a conocerse como Serial Endosymbiosis Theory (SET) (Teoría de la endosimbiosis seriada) en la que describe con concreción, mediante procesos simbiogenéticos, los pasos seguidos por las procariotas hasta la eclosión de las diferentes células eucariotas. Los tres pasos descritos por Margulis son:
Primera incorporación simbiogenética:
Una bacteria consumidora de azufre, que utilizaba el azufre y el calor como fuente de energía (arquea fermentadora o termoacidófila), se habría fusionado con una bacteria nadadora (espiroqueta) habiendo pasado a formar un nuevo organismo y sumaría sus características iniciales de forma sinérgica (en la que el resultado de la incorporación de dos o más unidades adquiere mayor valor que la suma de sus componentes). El resultado sería el primer eucarionte (unicelular eucariota) y ancestro único de todos los pluricelulares. El núcleoplasma de la células de animales, plantas y hongos sería el resultado de la unión de estas dos bacterias. A las características iniciales de ambas células se le sumaría una nueva morfología más compleja con una nueva y llamativa resistencia al intercambio genético horizontal. El ADN quedaría confinado en un núcleo interno separado del resto de la célula por una membrana.5
Segunda incorporación simbiogenética:
Este nuevo organismo todavía era anaeróbico, incapaz de metabolizar el oxígeno, ya que este gas suponía un veneno para él, por lo que viviría en medios donde este oxígeno, cada vez más presente, fuese escaso. En este punto, una nueva incorporación dotaría a este primigenio eucarionte de la capacidad para metabolizar oxígeno. Este nuevo endosombionte, originariamente bacteria respiradora de oxígeno de vida libre, se convertiría en las actuales mitocondrias y peroxisomas presentes en las células eucariotas de los pluricelulares, posibilitando su éxito en un medio rico en oxígeno como ha llegado a convertirse el planeta Tierra. Los animales y hongos somos el resultado de esta segunda incorporación.6
Tercera incorporación simbiogenética:
Esta tercera incorporación originó el Reino vegetal, las recientemente adquiridas células respiradoras de oxígeno fagocitarían bacterias fotosintéticas y algunas de ellas, haciéndose resistentes, pasarían a formar parte del organismo, originando a su vez un nuevo organismo capaz de sintetizar la energía procedente del Sol. Estos nuevos pluricelulares, las plantas, con su éxito, contribuyeron y contribuyen al éxito de animales y hongos.7
El primer paso, al día de hoy, no se considera demostrado. A finales de los años ochenta y principio de los noventa diversos trabajos no admitían las homologías propuestas entre los flagelos de los eucariontes y de las espiroquetas.8 9 10 11 Margulis defiende que las asociaciones entre espiroquetas y protistas apoyan su teoría, y "la comparación de genes y genomas arqueobaterianos con secuencias de eucariontes han demostrado la relación filogenética de ambos grupos".
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarLa Célula Bacteriana
Las bacterias son células muy sencillas; carecen de núcleo y tampoco presentan orgánulos en el citoplasma. Se las denomina Procariotas. Son organismos unicelulares y se encuentran en todos los ecosistemas.
Las bacterias son un numeroso grupo de seres vivos, con características muy diversas. En la clasificación de los Dominios, Woese, aparecen dos grupos de Procariotas, el Dominio Archaea, que engloba a los organismos más antiguos del Planeta, y el Dominio Bacteria, en el que se encuentran la gran mayoría de los organismos bacterianos actuales, también conocidos con el nombre de Eubacterias.
Las formas que presentan las bacterias pueden ser:
Coco
Bacilo
Vibrión
Espirilo
Las bacterias pueden presentarse como individuos sueltos, o formando colonias. Se pueden encontrar colonias de diplococos (bacterias redondeadas, de dos en dos), diplobacilos (bacterias alargadas, de dos en dos), estreptococos(cordones de bacterias redondeadas), estafilococos (masas laminares de bacterias redondeadas) o sarcinas(conglomerados tridimensonales de bacterias redondeadas).
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarLa Pared Celular En Gram + Y Gram –
BACTERIA GRAM POSITIVA
Tiene una capa gruesa de Peptidoglicano (mureina) y dos clases de ácidos teicoicos: Ácido Lipoteicoico que está en la superficie, empotrado en la capa de peptidoglicano y unido a la membrana citoplásmica. Y Ácido teicoico de la pared que está en la superficie y se une a la capa de peptidoglicano.
BACTERIA GRAM NEGATIVA
Tiene una capa delgada de Peptidoglicano (mureina) unida a una membrana exterior por lipoproteínas. La membrana exterior está hecha de proteína, fosfolípido y lipopolisacárido. En el lipopolisacárido, la porción de lípido está embebida en el fosfolípido y el antígeno O polisacárido entero se llama Endotoxina. La pared de la célula tiene poros llamados Porines para el transporte de substancias de peso molecular bajo.
Entre la membrana citoplásmica y la pared celular hay un espacio periplásmico con enzimas inactivadoras de antibióticos y proteínas de transporte.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarColoración De Gram
Es un tipo de tinción diferencial empleado en microbiología para la visualización de bacterias, sobre todo en muestras clínicas. Debe su nombre al bacteriólogo danés Christian Gram, que desarrolló la técnica en 1884. Se utiliza tanto para poder referirse a la morfología celular bacteriana como para poder realizar una primera aproximación a la diferenciación bacteriana, considerándose Bacteria Gram positiva a las bacterias que se visualizan de color violeta y Bacteria Gram negativa a las que se visualizan de color rosa.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarRibosomas Bacterianos
Son partículas constituidas por RNA y proteínas. El ribosoma bacteriano 70s consta de dos subunidades, pero es más pequeño que el de las células eucariotas. Es el lugar de síntesis de las proteínas, y el gran número de ribosomas refleja la importancia de esta función en la célula bacteriana. Se han encontrado algunas diferencias funcionales entre los ribosomas de las bacterias y los de las células eucariotas, tales diferencias han permitido encontrar compuestos con actividad antibiótica que actúan sobre el ribosoma bacteriano y no sobre el humano.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarFormas Y Agrupaciones Bacterianas
Los principales tipos de formas bacterianas son:
- cocos (células más o menos esféricas); - bacilos (en forma de bastón, alargados),
que a su vez pueden tener varios aspectos:
- cilíndricos
- fusiformes
- en forma de maza, etc.
Atendiendo a los tipos de extremos, éstos pueden ser:
- redondeados (lo más frecuente)
- cuadrados
- biselados
- afilados
espirilos: al igual que los bacilos, tienen un eje más largo que otro, pero dicho eje no es recto, sino que sigue una forma de espiral, con una o más de una vuelta de hélice.
vibrios: proyectada su imagen sobre el plano tienen forma de coma, pero en el espacio suelen corresponder a una forma espiral con menos de una vuelta de hélice.
Otros tipos de formas
filamentos, ramificados o no
anillos casi cerrados
formas con prolongaciones (con prostecas).
Estos distintos tipos de morfologías celulares deben de haberse originado por mecanismos evolutivos, a saber, por selección y estabilización adaptativa frente a las distintas presiones ambientales presentes en diferentes nichos ecológicos.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarFLAGELOS
Son filamentos proteicos responsables de la movilidad. Están constituidos por una proteína llamada flagelina. Se introducen en la membrana celular y se proyectan hacia fuera de la célula.
Los flagelos se disponen de distinta manera alrededor del cuerpo bacteriano. En algunas especies se encuentran en gran número y distribuídos sobre toda la superficie celular, a esta disposición se la llama peritrica. Cuando la bacteria tiene un único flagelo, en uno de los polos se denominamonotrica. Otras presentan un haz de flagelos en uno de los polos, se las llama lofotrica, las que presentan haces de flagelos en ambos polos, se las llama anfitricas, y las especies que carecen de flagelos se las denomina atricas.
Los flagelos mueven a la célula mediante rotación y le permiten a la bacteria responder a nutrientes químicos o materiales tóxicos y acercarse o alejarse de ellos por el fenómeno llamado quimiotáxis.
Ciertas bacterias usan otros medios de locomoción, las espiroquetas se mueven pero no poseen flagelos, presentan un filamento axial, que les permite el movimiento. Otras como las Mixobacterias, lo hacen por deslizamiento.
Esporas
Algunas especies de bacterias Gram positiva, como las de los géneros Bacillus, Clostridium y Sporosarcina , disponen de un notable mecanismo adaptativo cuando se ven sometidas a privación de nutrientes en su medio ambiente: si los niveles de fuentes de C, N, o P caen por debajo de un umbral, esto constituye una señal a la célula de que se avecina un largo período de privación de nutrientes. Entonces, la célula se implica en una serie de complejos cambios genéticos, metabólicos, y estructurales, proceso de esporulación o esporogénesis, que conducen a la diferenciación, en el interior de la célula vegetativa original, de una célula latente, que incluye la región nuclear y algo de citoplasma, separado por una pared de estructura compleja, del resto de la célula madre, se forma así la endospora, que es capaz de permanecer en estado durmiente, varios decenios, incluso siglos. Las esporas son fácilmente diseminadas por el aire; cuando caen en medios ricos en nutrientes, se desencadena su germinación, se reinicia la actividad metabólica, de modo que cada espora genera una nueva célula vegetativa, capaz de dividirse y llevar a cabo una actividad metabólica normal. O sea, la esporulación se puede considerar como un potente proceso de supervivencia de ciertas bacterias Gram positivas cuando se enfrentan a condiciones severas de hambre de nutrientes.
Las endosporas son formas de reposo (y no formas reproductivas), que representan una etapa del ciclo de vida de ciertas bacterias, y que se caracterizan por una estructura peculiar, diferenciada respecto de las células vegetativas, por un estado metabólico prácticamente detenido, y por una elevada resistencia a agentes agresivos ambientales, como desinfectantes químicos, radiaciones, el calor y la desecación.
La posición de la bacteria puede ser central, terminal o subterminal; y puede ser del mismo tamaño que la célula bacteriana, mayor o menor. En el género Bacillus, la espora es de igual tamaño que la célula bactriana, en el género Clostridium es de mayor tamaño y deforma el cuerpo bacteriano.
Gránulos de reserva/inclusiones citoplasmáticas
Son acúmulos de sustancias orgánicas o inorgánicas que se originan dentro del citoplasma bajo determinadas condiciones de crecimiento. Su presencia y cantidad depende de la especie bacteriana, del nivel de actividad metabólica y de la composición del medio de cultivo. Los gránulos pueden ser de glucógeno, de lípidos, de volutita, que son reservorios de energía en forma de metafosfatos, de azufre, de proteínas, etc.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarBacterias Patógenas
Las bacterias patógenas son una de las principales causas de las enfermedades y de la mortalidad humana, causando infecciones tales como el tétanos, la fiebre tifoidea, la difteria, la sífilis, el cólera, intoxicaciones alimentarias, la lepra y la tuberculosis. Hay casos en los que la etiología o causa de una enfermedad conocida se descubre solamente después de muchos años, como fue el caso de la úlcera péptica y Helicobacter pylori. Las enfermedades bacterianas son también importantes en la agricultura y en la ganadería, donde existen multitud de enfermedades como por ejemplo la mancha de la hoja, la plaga de fuego, laenfermedad de Johne, la mastitis, la salmonela y el carbunco.
Cada especie de patógeno tiene un espectro característico de interacciones con sus huéspedes humanos. Algunos organismos, tales como Staphylococcus o Streptococcus, pueden causar infecciones de la piel, pulmonía, meningitis e incluso sepsis, una respuesta inflamatoria sistémica que produce shock, vasodilatación masiva y muerte.[120] Sin embargo, estos organismos son también parte de la flora humana normal y se encuentran generalmente en la piel o en la nariz sin causar ninguna enfermedad.
Otros organismos causan invariablemente enfermedades en los seres humanos. Por ejemplo, el género Rickettsia, que son parásitos intracelulares obligados capaces de crecer y reproducirse solamente dentro de las células de otros organismos. Una especie de Rickettsia causa el tifus, mientras que otra ocasiona la fiebre de las Montañas Rocosas. Chlamydiae, otro filo de parásitos obligados intracelulares, contiene especies que causan neumonía, infecciones urinarias y pueden estar implicadas en enfermedades cardíacas coronarias.[121] Finalmente, ciertas especies tales como Pseudomonas aeruginosa, Burkholderia cenocepacia y Mycobacterium avium son patógenos oportunistas y causan enfermedades principalmente en las personas que sufren inmunosupresión o fibrosis quística.
Las infecciones bacterianas se pueden tratar con antibióticos, que se clasifican como bactericidas, si matan bacterias, o como bacterioestáticos, si solo detienen el crecimiento bacteriano. Existen muchos tipos de antibióticos y cada tipo inhibe un proceso que difiere en el patógeno con respecto al huésped. Ejemplos de antibióticos de toxicidad selectiva son el cloranfenicol y la puromicina, que inhiben el ribosoma bacteriano, pero no el ribosoma eucariota que es estructuralmente diferente.[124] Los antibióticos se utilizan para tratar enfermedades humanas y en la ganadería intensiva para promover el crecimiento animal. Esto último puede contribuir al rápido desarrollo de la resistencia antibiótica de las poblaciones bacterianas.[125] Las infecciones se pueden prevenir con medidas antisépticas tales como la esterilización de la piel antes de las inyecciones y con el cuidado apropiado de los catéteres. Los instrumentos quirúrgicos y dentales también son esterilizados para prevenir la contaminación e infección por bacterias. Los desinfectantes tales como la lejía se utilizan para matar bacterias u otros patógenos que se depositan sobre las superficies y así prevenir la contaminación y reducir el riesgo de infección.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarFORMAS L
Una Forma L es una variante bacteriana carente de pared celular (PC) o con PC defectuosa. Se estudia por primera vez en Streptobacillus moniliformis, donde se observa que produce en forma espontánea una serie de variantes capaces de reproducirse en forma de pequeños elementos filtrables carentes de PC o con PC defectuosa.
Estos microorganismos reciben el nombre de Formas L (L: proviene de Lister Institute, Londres). Estas bacterias tienen una morfología colonial muy parecida a la de los Micoplasmas.
Las Formas L se pueden presentar también en otras bacterias, siempre que la síntesis de PC esté alterada.
Formación
La formación de las Formas L, es favorecida por acción de la Penicilina, por enzimas líticas que digieren el peptidoglicano o por concentraciones elevadas de sales como ocurre con la mayoría de las bacterias Grampositivas. Algunas bacterias Gramnegativas pueden convertirse a Formas L en medios con osmolaridad normal o fisiológica.
Las Formas L son el equivalente morfológico de los Protoplastos y Esferoplastos, aunque el término de Formas L se limita a los organismos capaces de multiplicarse. Algunas Formas L, si se elimina el agente inductor, pueden revertir a la forma original (variante transitoria). Otras son Formas L estables.
Durante muchos años se creyó que las Formas L eran Micoplasmas, pero se diferencian de ellos porque:
a) Algunas Formas L pueden volver a poseer PC (formas inestables). Los Micoplasmas nunca poseen PC.
b) Las membranas celulares de las Formas L carecen de esteroles y/o carotenoides, a diferencia de lo que ocurre con los Micoplasmas en los que estas sustancias brindan rigidez a su membrana celular.
c) El análisis de homología de ácidos nucleicos demuestra que estos dos grupos no tienen ninguna vinculación taxonomica. La similitud de sus colonias y el parecido morfológico obedece a la ausencia de PC.
d) Las Formas L de algunas especies bacterianas, requieren una concentración de sales en el medio que funcione como estabilizador osmótico, para mantener la integridad celular.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarProtoplasto
Protoplasto, del griego antiguo πρωτον (primero) + πλασσειν (moldear), inicialmente la palabra se refiere al primer cuerpo organizado de una especie.
En biología un protoplasto es una célula de planta, bacteria u hongo que ha perdido total o parcialmente su pared celular, para lo cual se usan mecanismos enzimáticos. En el caso de eliminar la pared con enzimas se obtienen:
• Protoplastos - Cuando se elimina totalmente la pared celular.
• Esferoplastos - Cuando sólo se elimina parcialmente la pared.
Los protoplastos se obtienen a partir de bacterias Gram positiva y los esferoplastos de bacterias Gram negativa.
Esferoplasto
Un esferoplasto es una célula bacteriana desprovista de la mayor parte de su envoltura celular bacteriana mediante la adición de una penicilina. Los esferoplastos son muy frágiles y sensibles al estrés osmótico; tanto es así que se produce su lisis celular al transferirlos a un medio hipotónico.
Los esferoplastos de algunas bacterias Gram negativas han sido empleados en investigación científica para dilucidar los mecanismos de acción de loscanales iónicos de su membrana biológica mediante la técnica del patch clamp, originalmente diseñada para estudiar la excitabilidad de células nerviosas como las neuronas. Para el análisis de los esferoplastos bacterianos, se crecen las células en presencia de inhibidores de la división celular, de modo que crezcan pero que no se dividan. Esto acentúa notablemente su tamaño y modifica su forma hasta estructuras bacilares. En este punto, se añade la penicilina, lo que provoca el debilitamiento de la pared y el colapso de la estructura hasta una forma esférica más estable (con menor proporción superficie/volumen), forma cómodamente analizable mediante patch-clamp. Es común que la bacteria sometida a este proceso haya sido modificada genéticamente añadiendo un gen para un canal iónico sobreexpresado, lo que aumenta su abundancia y facilita su análisis. Esta técnica ha sido empleada E. coli a fin de estudiar los canales mecanosensitivos (MscL, MscS, and MscM) desde 1987. El sistema se ha extendido para el análisis de canales heterólogos; los esferoplastos gigantes de E. coli son un modelo de expresión comparable al del oocito de anfibio Xenopus
También es posible crear esferoplastos de levaduras como Saccharomyces cerevisiae mediante el empleo de enzimas como la zymolasa.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarNutrición Bacteriana
La nutrición es el proceso por el que los seres vivos toman el medio donde habitan las especies químicas que necesitanpara crecer.
Según el tipo de nutrición las bacterias se clasifican en:
1- Autótrofas o Fotótrofas, son aquellas especies de bacterias coloreadas( verdes, pardas, rojas, purpúreas) que al poseer pigmentos fotosintétricos como la Bacterioclorofila, bacterioviridina, etc, son capáces de trasnformar sustancias inorgánicas simples como agua, fotones de luz solar, CO2 en alimentos orgánicos por Fotosíntesis.
2- Quimiosintéticas, son aquellas especies de bacterias que obtiene la energía química ( ATP ) a partir de la Oxidación de sustratos inorgánicos como agua, óxidos, hidróxidos, ácidos, sales minerales, entre ellas las mas conocidas son las bacterias fijadoras de N2 atmosférico como Nitrosomonas, Nitrobácter, otras como las bacterias del Azufre, Desulfovibrio sp que por quimiosíntesis obtienen el ATP a partir de la oxidación de compuestos azufrados, luego excretan una vaina azufrada que e ubica sobre la pared celular.
3- Heterótrofas, son aquellas bacterias que obtiene la energía química a partir de la incorporación o Absorción directa de alimentos sintetizados por otros seres vivos, el Heterotrofismo bacteriano puede ser:
a- De tipo Parásito, en la cuál las bacterias obtiene los nutrientes del organismo del cuál hospedan o parasitan.
b- De tipo Saprófita, como la bacterias saprófitas que forman el eslabón de los Descomponedores en cadenas y redes tróficas, obtiene la energía química a partir de la descomposición de restos de organismos vegetales y animales en descomposición, entre ellas algunas especies mas conocidas están el Streptococus sanguis que por Saprofitismo de restos de alimentos que quedan entre los dientes origina la Placa bacteriana y los ácidos orgánicos son vertidos en el esmalte de los dientes causando las Caries dentales.
c- De tipo Sombiótica, como la especie Eschrichia Coli que habita en el tracto intestinal y que obtiene los nutrientes en condiciones normales del sistema digestivo del hombre y de todos los mamíferos pero por compensación de los nutrientes tomados en forma directa provee o sintetiza al organismo "vitamina K" que actúa como anticoagulante evitando la coagulación de la sangre dentro de los vasos sanguíneos.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarRespiración Bacteriana
Es un proceso complejo, que puede darse aeróbica o anaeróbicamente dependiendo de la bacteria, se acompaña con la liberación de energía la cual servirá para la síntesis de compuestos.
En las Bacterias la respiración puede ser de tipo:
- Aerobia
- Anaerobia Facultativa
- Anaerobia obligada.
En las bacterias Aerobias la respiración celular se realiza en la Cadena oxidativa o respiratoria asociada con la membrana plasmática que en ciertos casos se invagina formando una estructura llamada Mesosoma. A nivel de los Oxisomas localizados en el Mesosoma ya que carecen de mitocondrias los nutrientes son oxidados aeróbicamente liberando energía química y CO2.
En las Bacterias Anaerobias Facultativas (pueden o no fijar el O2 atmosférico) al respiración con O2 se realiza en los Oxisomas que forman las unidades fundamentales de la cadena oxidativa, utilizan la Glucólisis y luego la oxidación aerobia del ácido pirúvico en ATP y CO2, en tanto que aquellas bacterias que no fijan el O2 atmosférico la respiración es de tipo Anaerobia ( Fermentación) oxidando la Glucosa en condiciones anaerobias.
En las bacterias Anaerobias obligadas no poseen mecanismos para fijar el O2 atmosférico, por lo tanto la respiración en ellas es Anaerobia ( Fermentación).
Oxidan biológicamente la Glucosa por Glucólisis y luego trasnforman anaeróbicamente por Fermentación el ácido pirúvico en Etanal y posteriormente en Etanol (fermentación alcohólica).
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarTransmisión del material genético
Macho y hembra
Los testículos del toro y los ovarios de la vaca producen las células reproductoras por una serie de divisiones celulares que separan el número de cromosomas en una célula. El espermatozoide y el ovario contienen solamente un miembro del par de cromosomas.
Por lo tanto, las células de la vaca y del toro contienen 60 cromosomas (2n = 60), pero el espermatozoide en el semen y el óvulo en los ovarios contienen solamente 30 cromosomas (n=30, Figura 2). Los dos principios básicos de la transmisión de un rasgo (ejemplo sexo) son los siguientes:
1) Separación de los pares de cromosomas durante la formación de las células reproductoras;
2) Unión del espermatozoide con el óvulo para crear una nueva célula con un conjunto único de cromosomas.
Para 29 pares de cromosomas, ambos miembros son visualmente idénticos. De todas formas, para uno de los pares, un miembro es mucho más largo; es llamado cromosoma X, y el miembro más corto es llamado cromosoma Y. Todos los óvulos llevan el cromosoma X, pero el espermatozoide puede llevar ya sea el cromosoma X o el Y.
Rasgos cualitativos
Los rasgos cualitativos tienden a caer dentro de categorías discretas. Generalmente solo uno o unos pocos genes poseen un gran efecto sobre los rasgos cualitativos. El medio ambiente tene generalmente un pequeño papel al influenciar la categoría dentro de la que el animal cae. En este caso, el fenotipo de un animal refleja su genotipo. Ejemplos de rasgos cualitativos son:
* Color de pelo.
* Defectos hereditarios como enanismo.
* Presencia o ausencia de cuernos.
* Tipo sanguíneo.
Rasgos cuantitativos
Los rasgos cuantitativos difieren de los cualitativos de dos formas importantes:
1) Se encuentran influenciados por muchos pares de genes.
2) La expresión fenotípica es influenciada más fuertemente por el medio ambiente que en el caso de los rasgos cualitativos.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarAntibiograma
El antibiograma es la prueba microbiológica que se realiza para determinar la susceptibilidad (sensibilidad o resistencia) de unabacteria a un grupo de antibióticos. Las técnicas de antibiograma son las utilizadas en el laboratorio de microbiología para estudiar la actividad de los antimicrobianos frente a los microorganismos responsables de las infecciones.
Se considera como antimicrobiano cualquier sustancia con capacidad de matar o al menos de inhibir el crecimiento de los microorganismos y que sea susceptible de utilización como tratamiento en los pacientes. Pueden ser naturales, sintéticos o semisintéticos (modificación química de un compuesto natural). La historia moderna de los antibióticos comienza con el descubrimiento de sustancias presentes en unos microorganismos capaces de matar a otros microorganismos. La utilización de antibióticos supuso un avance enorme en la esperanza de vida de las personas que padecías procesos infecciosos, aunque desgraciadamente también supuso un aumento en los niveles de resistencia antibiótica.
El antibiograma tiene cuatro utilidades principales:
La utilidad básica del antibiograma es la instauración de un tratamiento antibiótico correcto al paciente. Es necesario conocer si el microorganismo responsable de la infección posee mecanismos que le confieran inmunidad frente a algún antibiótico para no incluirlo como terapia.
En cuanto al tratamiento el antibiograma no sólo es necesario en la instauración, también resulta útil en el seguimiento e incluso en la confirmación de tratamientos empíricos. En ocasiones la enfermedad infecciosa resulta grave y se comienza el tratamiento antes de conocer los datos de sensibilidad de la cepa. El antibiograma tiene que confirmar, o en su caso corregir el tratamiento.
Otra aplicación de las técnicas de estudio de resistencia es la epidemiología. Es necesario detectar el aumento de los niveles de resistencia en los aislamientos clínicos para tomar medidas correctoras.
Por otro lado también puede tener utilidad diagnóstica porque el perfil de resistencia puede en algún caso orientar en la identificación bacteriana.
Existen multitud de antimicrobianos en el mercado y el número sigue creciendo porque el desarrollo de más mecanismos de resistencia por parte de los microorganismos se traduce en un esfuerzo mayor por fabricar más y mejores antibióticos. No es necesario contrastar la eficacia de todos los antibióticos para cada aislamiento bacteriano. Los criterios que se siguen para seleccionar que antimicrobianos se ensayan respondes a factores de diversa índole:
Factores microbiológicos: Tipo de agente infeccioso y mecanismos de resistencia descritos previamente en su especie.
Factores farmacológicos: Tipo de antimicrobiano y parámetros de absorción, distribución y eliminación.
Factores del paciente: Tipo de infección. Factores de riesgo y estado general de salud. Situación inmunológica e hipersensibilidad.
Experiencia anterior referente a los patrones de resistencia antibiótica más habituales para cada especie.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarMecanismos De La Resistencia Bacteriana
La resistencia bacteriana es un fenómeno creciente caracterizado por una refractariedad
parcial o total de los microorganismos al efecto del antibiótico generado principalmente por
el uso indiscriminado e irracional de éstos y no sólo por la presión evolutiva que se ejerce
en el uso terapéutico.
El fenómeno de resistencia tiene un sustrato genético intrínseco o adquirido que se expresa
fenotípicamente por mecanismos bioquímicos. De esta manera puede observarse la
resistencia desde el ambiente biológico y otro el bioquímico.
Se conoce como resistencia natural a los mecanismos permanentes determinados
genéticamente, no correlacionables con el incremento de dosis del antibiótico. Un ejemplo
de esto es la resistencia de la Pseudomonas aeruginosa. a las bencilpenicilinas y al
trimetoprin sulfametoxazol; bacilos gram negativos aeróbicos a clindamicina.
La resistencia adquirida aparece por cambios puntuales en el DNA (mutación) o por la
adquisición de éste (plásmidos, trasposones, integrones).
En el primero se dan casos tales como la transformación de una Betalactamasa en una
Betalactamasa de espectro extendido o como en el caso de mutaciones de los genes que
codifican las porinas con el consecuente bloqueo del ingreso del antibiótico al interior del
microorganismo.
Existen otras denominaciones de resistencia como son:
• Resistencia relativa o intermedia: ocurre un incremento gradual de la MIC
(concentración inhibitoria mínima) a través del tiempo. Para obtener un efectoterapéutico es necesario alcanzar niveles séricos y tisulares adecuados. La susceptibilidad o resistencia del germen es en este caso dependiente de concentración.
• Resistencia absoluta: sucede un incremento súbito en la MIC de un cultivo durante o
después de la terapia. Es inefectivo el incremento de la dosis clínica usual. Ejemplo de
ello es la Pseudomonas spp. resistente a gentamicina y el Streptococcus pneumoniae
altamente resistente a penicilina y uso de levofloxacina.
• Seudorresistencia: ocurre una resistencia in vitro pero una gran efectividad in vivo.
Se denomina tolerancia antibiótica al fenómeno en el cual la diferencia entre la MBC
(concentración bactericida mínima) y la MIC es muy grande lo cual ocurre con relaciones
MBC/MIC mayores de 8 lo que permite la persistencia del microorganismo.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarVIROLOGÍA
La virología (rama de la microbiologia) es el estudio de los virus: su estructura, clasificación y evolución, su manera de infectar y aprovecharse de las células huésped para la reproducción del virus, su interacción con los organismos huéspedes, su inmunidad, la enfermedad que causan, las técnicas para su aislamiento, cultivo y su uso en investigación y terapia. La virología es considerada un sub-campo de la microbiología y la medicina. La virologia también es una rama taxonómica de la biología para estudiar los virus y todos sus componentes.
Su investigación incluye:
• la replicación viral
• los patógenos virales.
• la inmunología viral.
• las vacunas virales.
• los métodos de diagnóstico.
• la quimioterapia antiviral.
• las medidas de control de una infección.
• los diferentes signos que manifiestan la presencia de virus.
El Ciclo De Replicación De Los Virus consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden sobrevivir por si mismos en el exterior (véase Virus). Para reproducirse se introducirán en células donde controlarán sus mecanismos reproductivos.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarLa estructura de los virus.
La capsida, esta contiene una capa de proteína que la envuelve alrededor de un núcleo central de un producto químico altamente complejo llamado ácido nucleico, utilizado por el virus para su reproducción. Típicamente, la capsida se divide en subunidades llamadas los capsomeros. Las radiografias han mostrado que los virus tienen una capsida en forma de un sólido de 30 caras.
EL Cuerpo, posee una simetría compleja, asociada a la capsida hay un vástago con una estructura que consiste en una vaina retráctil que rodea a un núcleo y es usada a modo de inyección.
La cola, localizada al final del nucleo, es una placa espigada que lleva 6 fibras delgadas, que ayuden a asegurar al virus a sujetarse a la célula anfitrión, durante la invasión de la misma.
En una serie de experimentos realizados en 1955,por Fraenkel Conrat y Williams ,demostraron que el virus del mosaico del tabaco (TMV),se formaba espontáneamente cuando se mezclaba una proteína purificada de la capa y su RNA geonómico ,al ser incubadas juntas, la estructura del TMV se regenera por si sola. A pesar de la gran variabilidad mostrada en las características del virus, todos se basan en algunas características básicas.
Metodos de analisis
La microscopia electrónica ha sido muy útil en dar información acerca de los virus con gran resolución, en dicho instrumento el nivel de la resolución es de 5nm (un nanometro 1nm equivale a 10^ -9 metros es decir 1 dividido 1000 millones de metro, para dar una cierta clase de perspectiva a esta dimensión diremos que:
un átomo mide de 0,2-0,3 nm de diámetro
una hélice de una proteína tiene 1nm de diámetro
el ADN posee 2nm de diámetro
La imagen obtenida con el microscopio electrónico puede darnos la forma de un virus con gran resolución, sin embargo para aumentarla a la escala atómica conveniente utilizar la cristalografía de rayos X, esto requiere que un virus pueda ser cristalizado, la cristalización de un virus fue descubierta en los años 30 y la estructura se puede determinar utilizando un modelo de difracción revelado por esta técnica , la primera estructura con resolución atómica de un virus fue realizada en 1978 y desde entonces muchas estructuras virales se han determinado. En algunos casos las estructuras se determinan con cristales de virus y en otros casos esta técnica no es posible. Un análisis bioquímico se puede utilizar para determinarse que componentes están presentes en el virus y en que relación. Los componentes individuales del virus, se cristalizan y sus estructuras se determinan. La clonación del gene y las técnicas de secuenciados facilitan el aislamiento de grandes cantidades proteínas purificadas y de los ácidos nucleicos para este propósito. La estructura total entonces es deducida construyendo el virus con estas subunidades, teniendo en cuenta también la información de otras técnicas tales como microscopia electrónica.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarSIMETRÍA VIRAL
La simetría es la forma que adopta un virus en el espacio y está dad por la estructura de su nucleocápside. La simetría puede ser helicoidal, icosaédrica o compleja.
Los virus con simetría helicoidal presentan una nucleocápside cilindrica que puede estar extendida como en el virus del mosaico del tabaco. Este es un virus desnudo. En otros casos la nucleocápside está arrollada sobre sí misma y recubierta por una envoltura, como en el virusInfluenza.
Los virus con simetría icosaédrica son poliedros regulares con 20 caras triangulares, 30 aristas y 12 vértices. Son ejemplos de virus con simetría icosaédrica desnudos el virus de la poliomielitis o losadenovirus. Los herpesvirus y togavirus son virus con simetría icosaédrica y recubiertos por una envoltura.
Se denominan virus de simetría compleja a aquellos que presentan una nucleocápside helicoidal o icosaédrica recubierta por una envoltura laxa. Ejemplos de éstos virus son los poxvirus (forma de ladrillo), los rhabdovirus (forma de bala) y algunos bacteriófagos que poseen una cabeza con simetría icosaédrica y una cola con simetría helicoidal.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarFAMILIAS DE VIRUS
I- Adenoviridae
Mastadenovirus
Es un taxón que afecta a vertebrados.
Aden-(var.”adeno-“) Elemento prefijo del gr. “adén”, que significa “glándula”.
Viriones isométricos no envueltos.
Nucleocapside angular 70-90 nm diámetro.
Simetría icosaedrica.
II- Virus de la peste porcina africana
Infecta a cerdos domesicos, facoceros, potamóquero de río y garrapatas.
Causa fiebres hemorrágicas.
La peste africana se limitaba al continente que lleva su nombre hasta 1957 cuando la enfermedad se informó en otros países.
III- Arenaviridae
Virus que afecta a vertebrados.
Arena, viene del latín con la familia de términos del árido.
Virus envueltos, ligeramente pleomórficos; esféricas. Viriones probablemente contengan 2 nucleocapsides envueltas.
Causantes de fiebres hemorrágicas.
IV- Artevivirus
Familia de virus que infectan animales.
Arterivirus: arteritis. Inflamación vasos sanguíneos.
Estos virus son pequños, con envoltura y con forma icosaédrica.
V- Astroviridae
Virus que afecta a vertebrados.
Astro- es un prefijo lo cual significa estrellas.
Human astrovirus: Virones no envueltos. Nucleocapside isométrica. 27-30 nm diámetro. Simetría polihedrica. Con vértices.
VI- Baculoviridae
Virus que infecta vertebrados. Báculo: del latín, “palo”. Refiriéndose a la forma nucleocápside.
Son visibles en secciones de tejido infectado. Viriones ocluidos por cristales de proteínas o no ocluidos. Virion envuelto y pleomorfo forma de vara. Nucleocapsides en forma de vara. 200-450 nm long. Y 30-100 de diámetro.
VII- Badnavirus
Afecta a plantas
Etimología: Baciliforme Ba, DNA.
VIII- Barnaviridae
Infecta a hongos.
Etimología: Baciliforme Ba, RNA.
Sin envoltura, con un baciliformes T=1 simetría icosaédrica, aproximadamente 50 nm de longitud y 20 nm de gran tamaño.
IX- Birnaviridae
Afecta a vertebrados.
Birna: para el ARN bipartito.
No envueltos, de 60 nm de diámetro, virión icosaédricO con T=13 simetría.
X- Bromoviridae
Afecta a plantas.
Bromo- porque infecta a Bromus inermis.
Viriones no envueltos, icosaédrico o baciliformes 26-35 nm de diámetro.
XI- Bunyaviridae
Afecta a vertebrados y a plantas.
Viriones pleomorficos, esferoidales.
“Bunyamwera” ciudad de Uganda donde se aisló.
Envueltos, esféricos. Diámetro de 80 a 120 nm.
XII- Caliciviridae
Virus que infecta a vertebrados y a invertebrados.
“Calici” porque presenta depresiones calciformes.
Viriones no están envueltos y la capside es isométrica. Simetría icosaedrica.
XIII- Capillovirus
Infecta a plantas.
Del latín “capella”, diminutivo de capa.
El 3 es poliadenilado. Codifica por lo menos 3 proteínas.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarDIAGNÓSTICO DE LAS VIROSIS
Los virus son organismos tan pequeños, que su tamaño les permite pasar a través de los filtros que retienen bacterias. Las características básicas para identificar a un virus son: la composición del ácido nucleico, la disposición de la cápside, la presencia o ausencia de una envoltura, el tamaño del virus y el huésped.
DIAGNOSTICO VIRAL
1. Métodos para detectar virus.
1.1. Observación del efecto citopático, en cultivo de tejidos inoculados con ciertos virus. La etiología de un determinado síndrome viral puede demostrarse recuperando el agente causal en tejidos celulares de diversos tipos (por ej: riñón de mono, pulmón humano fetal, amnios humano o células tumorales humanas). Los huevos embrionados actualmente solo se utilizan para cultivar el virus cuando se desea preparar vacunas. El empleo de animales de experimentación es excepcional en el diagnóstico.
• Las células primarias de riñón de mono son excelentes para cultivar mixovirus, paramixovirus y muchos enterovirus; también para algunos adenovirus.
• Las células diploides fetales humanas son células fibroblásticas en las que crece un amplio espectro de virus, como el de herpes simple, varicela zoster, adenovirus, picornavirus, y sólo en éstas pueden cultivarse los citomegalovirus.
• Las células HEP-2 son epiteliales provenientes de un cancer humano, útiles para recuperar el virus respiratorio sincitial, adenovirus, y herpes simple.
• Células mononucleares normales normales humanas de sangre periférica, que no suelen estar disponibles, permiten el cultivo del virus ( SIDA ) de la inmunodeficiencia, el coxsackie A y los togavirus.
El crecimiento de un virus se detecta mediante la observación de cambios en la monocapa del cultivo celular, denominado efecto citopático (ECP). Cuando aparecen cambios en la morfología celular, lisis celular, vacuolización, formación de cincicios y existen cuerpos de inclusión, se produce un efecto citopático característico. Los cuerpos de inclusión son cambios histológicos que se producen en las células por los componentes virales o por variaciones en las estructuras celulares.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarContinuación desarrollo del Tema: Diagnóstico Viral
Algunos virus no producen rápidamente ECP en las líneas celulares que se suelen utilizar en los laboratorios de virología clínica. No obstante se pueden determinar por otras técnicas, como la interferencia en la replicación de otros virus (por ej los picornavirus no pueden replicarse en células previamente infectados con el virus de la rubeola, lo que se denomina interferencia heteróloga).
1.2. Fijación de eritrocitos en cultivos celulares infectados con virus. En el caso de virus que dan lugar a este fenómeno, como mixovirus o virus de la parotiditis, se observa una hemaglutinación al adicionar eritrocitos a la monocapa de células.
1.3. Detección de antígenos virales. Cuando los virus se pueden detectar antes de desarrollar el efecto citopático, se utilizan anticuerpos monoclonales dirigidos contra antígenos tempranos y técnicas de inmunofluorescencia o inmunoperoxidasa.
1.4. Inmunofluorescencia directa, utilizando células del paciente, como leucocitos, orina o mucosa oral o genital, para buscar virus.
1.5. Detección de ácidos nucleicos virales por reacción de polimerasa en cadena, las cuales permiten amplificar una porción pequeña de ácido nucleico en un período de tiempo corto.
1.6. Microscopía electrónica. Cuando algunos virus comunes no se aislan con los métodos de cultivo habituales, ni pueden diagnosticarse por técnicas serológicas, pueden estudiarse con otras técnicas, como la microscopía electrónica. Tal es el caso de los calcivirus, los minirrotavirus, los astrovirus y los parvovirus del cerebro. Sin embargo, hay tres grandes dificultades para su uso: el instrumento es caro y es difícil su mantenimiento. Por otro lado, deben estar presentes grandes cantidades de particulas virales en la muestra para poder detectarse. Finalmente, la morfología no es suficientemente diferencial para un diagnóstico preciso.
2. Métodos para detectar anticuerpos específicos virales.
La mera presencia de anticuerpos específicos en contra de un determinado virus, no permite el diagnóstico de dicha infección. Debe tomarse en cuenta el cambio en los títulos en un lapso corto de tiempo, o ver si hay seroconversión. En muchas virosis la presencia de IgM sugiere infección reciente.
2.1. Fijación de complemento: Es una técnica estándar, pero de difícil ejecución. La muestra se procesa con el antígeno o el anticuerpo y el complemento; el complemento residual se determina mediante la lisis de eritrocitos cubiertos de anticuerpos que actúan como indicadores. Los anticuerpos medidos por este método se suelen desarrollar tardíamente en el desarrollo de la enfermedad. Los miembros de algunos grupos como adenovirus, influenza A y B, poseen antígenos comunes que se detectan por este método. Los demás grupos deben analizarse individualmente.
2.2. Neutralización: Cuando se incuba un virus con anticuerpos homólogos específicos de tipo, el virus no es capaz de producir la infección en un sistema de cultivos celulares indicadores. La respuesta de anticuerpos neutralizantes es específica del tipo devirus y se desarrolla al comienzo de los síntomas, elevándose los títulos rápidamente durando mucho tiempo.
2.3. Inhibición de la hemaglutinación: Se puede realizar con diversos virus que aglutinan selectivamente los eritrocitos de varias especies animales (por ej: pollo, cobaya y humanos). La capacidad de hemaglutinación del suero se inhibe por el suero humano específico. Los anticuerpos inhibidores de la hemaglutinación se desarrollan tras el comienzo de los síntomas, posteriormente se elevan y al final disminuyen. Esta prueba es útil para detectar virus de rubeola y para determinar el estado inmunitario.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarETAPAS DE LA REPLICACIÓN VIRAL
1- El primer paso en la infección viral es la absorción a la membrana de la célula susceptible por medio de la adhesión de ligandos virales (proteínas de la capside o glucoproteínas de las espículas, por ejemplo a receptores superficiales de la célula). La distribución en el cuerpo del hospedador de ciertos tipos de receptores celulares explica el tropismo tisular y de hospedador de los virus. Los tejidos y células que carecen de receptores específicos de virus determinados no son infectados por dichos viriones. La naturaleza de los receptores es variable y ciertos virus pueden tener más de un tipo de receptor.
2- Los virus complejos producen una rotura en la membrana celular del hospedador en uno de los puntos de anclaje, gracias a la presencia de algunas moléculas de enzimas hidrolíticas entre lasproteínas de la cápsida. A través de la rotura, el tubo central inyecta el ADN vírico, quedando la cápside vacía en el exterior de la célula diana y el ácido nucleico libre en el citoplasma. La presencia de cápsidas en la superficie celular es un buen indicio de que ha sufrido una infección vírica.
Otros virus sin envoltura lipídica se introducen en la célula con cápsida y todo, lo cual puede realizarse de dos maneras:
• Por penetración directa: después de la fijación, el virus abre una brecha en la membrana y se introduce en el citoplasma.
• Por endocitosis: la membrana forma una invaginación en torno al virus, llegando a formar una vesícula que penetra en la célula. Formada la vesícula, el virus abre una brecha en la membrana de la misma con ayuda de algunas enzimas hidrolíticas que él mismo transporta, penetrando así en el citoplasma.
Los virus con envoltura lipídica burlan la barrera de la membrana celular porque su cubierta lipídica se funde con la membrana, ya que son de la misma naturaleza. Esta fusión de membranas puede realizarse en dos lugares distintos:
• Fusión en la superficie celular: de manera que el virión penetra directamente en el citoplasma.
• Fusión con un lisosoma: se forma una vesícula por endocitosis, a la que se une un lisosoma para digerir la partícula introducida; entonces, la cubierta lipídica del virus se funde con la membrana del lisosoma y el virión escapa hacia el citoplasma.
3- Según la duración de la fase de eclipse, se suelen distinguir dos modalidades de ciclo infeccioso de un virus:
• Ciclo ordinario: el ácido nucleico vírico procede inmediatamente a la transcripción de su mensaje genético en los ARN necesarios para su multiplicación, y prosigue rápidamente el ciclo vital. Este tipo de ciclo es el más extendido en la naturaleza.
• Ciclo lisogénico: fue descubierto por André Lwoff en bacteriófagos. El ADN vírico se cierra por sus extremos generando un ADN circular. Este ADN se inserta en el ADN bacteriano en un lugar específico en el que la secuencia de nucleótidos bacterianos es semejante alguna región del ADN vírico.
La bacteria prosigue sus funciones vitales sin que el virus realice ninguna acción, y cuando el ADN bacteriano se duplica también lo hace el ADN vírico, de manera que el genoma del virus pasa a las dos bacterias hijas. La multiplicación bacteriana puede seguir durante generaciones sin que el virus se manifieste. Pero ante una alteración de las condiciones ambientales, el ADN vírico se separa del bacteriano y prosigue entonces las restantes fases de ciclo infeccioso, produciendo la muerte de la bacteria y nuevos ejemplares del virus.
4- La multiplicación del virus consta de la replicación de su material genético, de la transcripción de su mensaje en una molécula de ARN y de la traducción del mensaje para producir proteínasvíricas, tanto las que formarán parte de la cápsida como las proteínas enzimáticas necesarias para el ensamblaje de las piezas del virión y para algunas de las funciones anteriores. Losribosomas y la mayor parte de las enzimas que los ácidos nucleicos víricos utilizan en estos procesos son los de la célula infectada.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarANTIVIRALES
Los antivirales son un tipo de medicamento usado para el tratamiento de infecciones producidas por virus. Tal como los antibióticos (específicos para bacteria), existen antivirales específicos para distintos tipos de virus. No sin excepciones, son relativamente inocuos para el huésped, por lo que su aplicación es relativamente segura. Deben distinguirse de los viricidas, que son compuestos químicos que destruyen las partículas virales presentes en el medio ambiente.
Descripción
Muchos de los antivirales disponibles actualmente están diseñados para ayudar el tratamiento del VIH (virus del sida), herpesvirus, productores de la varicela, el herpes labial, el herpes genital, etc. y los virus de la hepatitis B y C, que pueden causar cáncer de hígado. Los investigadores están trabajando actualmente para extender el rango de antivirales a otras familias de patógenos.
Características de un antiviral
Para que un fármaco antiviral sea considerado para el tratamiento de una infección viral debe cumplir ciertos requisitos indispensables. El fármaco debe ser específicamente activo contra el virus“target” (blanco), inhibiendo algún paso esencial de su metabolismo y debe poder debilitar a las cepas resistentes que puedan surgir. Además, debe tener ciertos parámetros que permitan al fármaco ser de alta biodisponibilidad oral, rápida penetración en los tejidos infectados y atóxicos para las células normales y el organismo en general a corto y largo plazo. El fármaco debe ser transformado a su forma activa solamente en células infectadas, debe ser excretado sin ser metabolizado o sin generar metabolitos tóxicos, y debe tener baja frecuencia de administración. Finalmente, el fármaco debe tener bajo costo para que pueda estar disponible para toda la población. Todas estas características, entre otras, dificultan el descubrimiento de nuevos fármacos efectivos para el tratamiento de las enfermedades virales.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarMecanismo De Acción De Los Retrovirales
Los fármacos antirretrovirales no matan al virus de la inmunodeficiencia humana (VIH). Ni siquiera logran dormirlo. Lo 'atontan' un poco y lo mantienen en un estado de reposo, pero que le permite, de vez en cuando, salir a 'pasear' por las células del organismo humano. Lo ha descubierto un equipo de investigadores españoles, que pone así fin a una de las controversias médicas más extendidas en el campo del sida.
Durante años, la teoría más apoyada ha sido la que consideraba que el cóctel de fármacos impedía la replicación del virus. No era capaz de eliminarlo, pero sí podía evitar que siguiera infectando. Otra hipótesis, más minoritaria, indicaba que a pesar de la terapia, el virus podía mantener algo de actividad. Y es esta última la que finalmente se ha confirmado, según la investigación que publica 'Nature Medicine' y que ha sido realizada por expertos del Instituto de Investigación del Sida IrsiCaixa, ubicado en el Hospital German Trias i Pujol de Badalona.
Lo que han visto los investigadores, dirigidos por Javier Martínez Picado, es que algunos pacientes infectados por el VIH, a pesar de que presentan una carga viral indetectable gracias al tratamiento y que parecen tener la infección controlada, cuentan con una pequeña proporción del virus que sigue siendo infeccioso. Un descubrimiento que, según sus autores, "tiene importantes implicaciones clínicas, ya que podría explicar por qué, a pesar de que los pacientes permanecen en terapia durante muchos años, los fármacos no permiten curar la infección de manera definitiva".
Martínez Picado lo aclara de forma sencilla a ELMUNDO.es: "Un paciente infectado sin tratamiento es como un bosque en llamas. Con los fármacos logramos apagar el fuego, pero quedan rescoldos y basta con un pequeño golpe de aire para que vuelva a incendiar. Esto es lo que hace el virus. Está apagado, pero no muerto".
Carga viral indetectable
Para llegar a estas conclusiones, el equipo analizó a 69 pacientes que seguían la terapia antirretroviral y que tenían carga viral indetectable (cantidad de virus en sangre) durante una media de cinco años. A 45 de ellos se les intensificó el tratamiento añadiendo otro fármaco, el Raltegravir, el primero de una nueva familia de medicamentos antisida, que bloquea la acción de la integrasa (una enzima que utiliza el VIH para copiar su material genético en el ADN de las células del paciente). Los otros participantes siguieron con su terapia antirretroviral normal.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarVIROIDES
Los viroides son agentes infecciosos que, al igual que los virus, tienen un ciclo extracelular que se caracteriza por la inactividad metabólica y unciclo intracelular en el que causan infección al huésped susceptible, pero que a diferencia de los virus, los viroides no poseen proteínas ni lípidos y están constituidos por una cadena cíclica corta de ARN, circular o con forma de varilla, (que no codifica proteínas).1 Es importante decir que tanto su forma intracelular como extracelular son las mismas (ARN desnudo), los mecanismos por los cuales éstos logran causar infección están relacionados con la autocatálisis de su material genético. En sí constituyen una etapa primitiva de los virus. El primer viroide fue descubierto por T. O. Diener en 1978, al intentar identificar el agente causal de una enfermedad que inicialmente supuso era inducida por un virus llamada "la enfermedad del tubérculo fusiforme de la patata" (potato spindle tuber o PSTVd) La nomenclatura de los viroides incluye el sufijo “Vd” para distinguirlos de los virus. El segundo viroide en ser descubierto fue el agente causal de la exocortis de los cítricos (citrus exocortis viroid, CEVd). Actualmente se conocen más de 200 viroides.
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarOtros Biones Acelulares
Satélites (Schneider): Virus asociados a otros virus de los que dependen para su multiplicación y capacidad para causar infección. Sólo de vegetales. Son incapaces de multiplicarse sin la concurrencia de un virus y controlan la capacidad de los virus. Actúan como parásitos de virus.
2- Virusoides o sateloides (Fransky): pequeñas hebras de ARN monocatenario circular, localizados dentro de otros virus ARN.
3- Elementos transponibles o transposones.
4- Plásmidos
Nombre: Samely Donastorg Mercedes. Matrícula: 2011-0544
ResponderEliminarPRIONES
Un prión es un agente infeccioso que se compone principalmente de proteínas. Hasta la fecha, todos los agentes que han sido descubiertos propagan por transmitir un Estado de mis-folded de proteínas; la proteína misma no máquinas y el proceso depende de la presencia del polipéptido en el organismo hospedador. La mis-folded forma de la proteína del prión ha sido implicada en una serie de enfermedades en una gran variedad de mamíferos, incluyendo encefalopatía espongiforme bovina (EEB, también conocida como "enfermedad de las vacas locas") en el ganado y la enfermedad Creutzfeldt - Jakob (ECJ) en los seres humanos. Todas las enfermedades priónicas conocidos afectan a la estructura del cerebro o de otros tejidos neuronales, y todos están actualmente intratable y siempre son fatales. En general uso de prión se refiere a la unidad teórica de la infección. En notación científica, PrPC se refiere a la forma endógena de la proteína del prión (PrP), que se encuentra en multitud de tejidos, mientras que PrPSc se refiere a la forma mal doblada de PrP, que es responsable de la formación de placas de amiloide y la neurodegeneración.
Priones son hipótesis a infectar y propagarse por células anormalmente en una estructura que es capaz de convertir las moléculas de la proteína normales en forma anormalmente estructurada. Todos conocido priones inducen la formación de un pliegue de amiloide, en los que la proteína polymerises en un agregado que consta de láminas beta estrechamente empaquetados. Esta estructura alterada es muy estable y se acumula en el tejido infectado, causando la muerte de células y daño del tejido. Esta estabilidad significa que los priones son resistentes a la desnaturalización por agentes químicos y físicos, dificultando la eliminación y contención de estas partículas.
Mostrando el comportamiento del prión-tipo de proteínas también se encuentran en algunos hongos, que ha sido útil para ayudar a comprender los priones mamíferos. Priones hongos, sin embargo, no parecen causar enfermedades a sus anfitriones y incluso pueden conferir una ventaja evolutiva a través de una forma de herencia basados en proteína.
La palabra prion es una palabra compuesta que se deriva de las letras iniciales de las palabras proteinaceous'' y menfectious'', con -en añadido por analogía a la palabra virien.
2011-0119 dios te bendiga
ResponderEliminarLista de arbovirus (incompleta)
Familia Togaviridae
El virus chikungunya (CHIKV) (Vector: Mosquito)
El virus O'nyong-nyong (Vector: Mosquito)
El virus del Río Ross (Vector: Mosquito)
El virus Sindbis (Vector: Mosquito)
Familia Flaviviridae[editar]
Fiebre amarilla (Vector: Mosquito)
Dengue (Vector: Mosquito)
Encefalitis japonesa (Vector: Mosquito)
Virus Kunjin (Vector: Mosquito)
Enfermedad de Kyasanur (Vector: Garrapata)
virus Usutu (USUV) (Vector: Mosquito)
Virus del Nilo Occidental (Vector: Mosquito)
Encefalitis de San Luis (Vector: Mosquito)
Familia Bunyaviridae
La Fiebre del Valle del Rift (Vector: Mosquito)
Encefalitis de La Crosse (Virus de La Cross) (Vector: Mosquito)
Fiebre por Orepuche (Virus de Orepuche) (Vector: Mosquito)
Virus Toscana (VTOS) (Fiebre Pappataci) (Vector: Mosquito)
Virus Trivittatus (Vector: Mosquito)
Familia Reoviridae
Lengua azul (Vector: Mosquito)
Fiebre del Colorado por garrapatas (Vector: Garrapata)
Virus Lipovnik (LIPV) (Vector: Garrapata)
Virus Tribec (Vector: Garrapata)
Virus Eyach (Vector: Garrapata)
Familia Asfarviridae
dios te bendiga 2011-0119 partes 4
ResponderEliminarMecanismo De Acción De Los Retrovirales
Los fármacos antirretrovirales no matan al virus de la inmunodeficiencia humana (VIH). Ni siquiera logran dormirlo. Lo 'atontan' un poco y lo mantienen en un estado de reposo, pero que le permite, de vez en cuando, salir a 'pasear' por las células del organismo humano. Lo ha descubierto un equipo de investigadores españoles, que pone así fin a una de las controversias médicas más extendidas en el campo del sida.
Durante años, la teoría más apoyada ha sido la que consideraba que el cóctel de fármacos impedía la replicación del virus. No era capaz de eliminarlo, pero sí podía evitar que siguiera infectando. Otra hipótesis, más minoritaria, indicaba que a pesar de la terapia, el virus podía mantener algo de actividad. Y es esta última la que finalmente se ha confirmado, según la investigación que publica 'Nature Medicine' y que ha sido realizada por expertos del Instituto de Investigación del Sida IrsiCaixa, ubicado en el Hospital German Trias i Pujol de Badalona.
Lo que han visto los investigadores, dirigidos por Javier Martínez Picado, es que algunos pacientes infectados por el VIH, a pesar de que presentan una carga viral indetectable gracias al tratamiento y que parecen tener la infección controlada, cuentan con una pequeña proporción del virus que sigue siendo infeccioso. Un descubrimiento que, según sus autores, "tiene importantes implicaciones clínicas, ya que podría explicar por qué, a pesar de que los pacientes permanecen en terapia durante muchos años, los fármacos no permiten curar la infección de manera definitiva".
Martínez Picado lo aclara de forma sencilla a ELMUNDO.es: "Un paciente infectado sin tratamiento es como un bosque en llamas. Con los fármacos logramos apagar el fuego, pero quedan rescoldos y basta con un pequeño golpe de aire para que vuelva a incendiar. Esto es lo que hace el virus. Está apagado, pero no muerto".
Carga viral indetectable
Para llegar a estas conclusiones, el equipo analizó a 69 pacientes que seguían la terapia antirretroviral y que tenían carga viral indetectable (cantidad de virus en sangre) durante una media de cinco años. A 45 de ellos se les intensificó el tratamiento añadiendo otro fármaco, el Raltegravir, el primero de una nueva familia de medicamentos antisida, que bloquea la acción de la integrasa (una enzima que utiliza el VIH para copiar su material genético en el ADN de las células del paciente). Los otros participantes siguieron con su terapia antirretroviral normal.
partes 1 y 2
ResponderEliminarLa virología (rama de la microbiologia) es el estudio de los virus: su estructura, clasificación y evolución, su manera de infectar y aprovecharse de las células huésped para la reproducción del virus, su interacción con los organismos huéspedes, su inmunidad, la enfermedad que causan, las técnicas para su aislamiento, cultivo y su uso en investigación y terapia. La virología es considerada un sub-campo de la microbiología y la medicina. La virologia también es una rama taxonómica de la biología para estudiar los virus y todos sus componentes.
ESTRUCTURA DE LOS VIRUS
En los virus se distinguen las partes siguientes:
1.- Genoma vírico. Se compone de una o varias moléculas de ADN o de ARN, pero nunca los dos simultaneamente, Se trata de una sola cadena, abierta o circular, monocatenaria o bicatenaria. .
2.- Cápsida. Es la cubierta proteica que envuelve al genoma vírico. Está formada por proteínas globulares o capsómeros que se disponen de una manera regular y simétrica, lo que determina la existencia de varios tipos de cápsidas: icosaédricas, helicoidales y complejas.
La función de la cápsida es proteger el genoma vírico y, en los virus carentes de membrana, el reconocimiento de los receptores de membrana de las células a las que el virus parasita.
3.-Envoltura membranosa. Formada por una doble capa de lípidos que procede de las células parasitadas y por glucoproteínas incluidas en ella cuya síntesis está controlada por el genoma vírico. Las glucoproteínas sobresalen ligeramente de la envoltura y tienen como función el reconocimiento de la célula huesped y la inducción de la penetración del virus en ella mediante fagocitosis.
evelyn mota 2011-0637
ResponderEliminarDIAPOSITIVA # 3
LA CÉLULA BACTERIANA
Las bacterias son células muy sencillas; carecen de núcleo y tampoco presentan orgánulos en el citoplasma. Se las denomina Procariotas. Son organismos unicelulares y se encuentran en todos los ecosistemas.
Las bacterias son un numeroso grupo de seres vivos, con características muy diversas. En la clasificación de los Dominios, Woese, aparecen dos grupos de Procariotas:
el Dominio Archaea, que engloba a los organismos más antiguos del Planeta, y
el Dominio Bacteria, en el que se encuentran la gran mayoría de los organismos bacterianos actuales, también conocidos con el nombre de Eubacterias.
Las bacterias se caracterizan por no tener núcleo patente, ni clorofila típica, aunque pueden poseer otros pigmentos similares como bacterioclorina, bacterioclorofila y bacteriopurpurina, y sin pseudópodos. La bacterioclorina es un pigmento verde fotosintetizante que, según ciertos autores, acompaña a la bacteriopurpurina y a la bacterioclorifila en las algas purpúreas. La bacterioclorofila es otro pigmento fotosintetizante que, junto con la bacteriopururina, poseen las bacterias purpúreas. La bacteriopurpurina es un pigmento rojo que enmarca a la bacterioclorifila en las bacterias purpúreas, y que colabora con ella en la síntesis de los glúcidos, a partir del bióxido de carbono atmosférico. Esta síntesis se diferencia de la clorofílica en que utiliza el ácido sulfuhídrico como donador de hidrógeno.
Aunque la organización de las bacterias es de tipo celular, el microscopio electrónico ha puesto al descubierto numerosas características propias de la célula bacteriana. También pueden ser observadas con el microscopio óptico.
Una célula bacteriana se compone de una pared celular, membrana, citoplasma y ácido nucleico. La pared bacteriana aísla y protege perfectamente a la bacteria. Incluso algunas bacterias tienen una cápsula externa que las protege de los antibióticos y de los anticuerpos. La membrana bacteriana es esencialmente idéntica a la de las células eucariónticas, aunque posee unos entrantes en el citoplasma. En el citoplasma bacteriano las únicas estructuras existentes son los ribosomas y algunas vesículas llenas de gas. El ácido nucleico está formado por una sola cadena de ADN, que se suele llamar cromosoma bacteriano y es de forma circular, que se diferencia del cromosoma eucariótico en que es más pequeño y no se asocia tan íntimamente con las proteínas. Ambos se parecen en que se componen de ADN. Éste se halla condensado en una región de la bacteria llamada nucleoide o falso núcleo.
Sus dimensiones son muy reducidas: tan sólo de algunas micras o fracción de micra. Se pensaba, no hace aún muchos años, que fuesen el límite inferior en la escala de ser vivientes; pero los modernos microscopios permiten observar otros seres mucho más diminutos, tales las rickettsias y los virus.
Unas bacterias son inmóviles, otras poseen minúsculos flagelos, cuyo número y distribución varía notablemente, que les permiten desplazarse.
Su capacidad reproductora es enorme, pues algunas se dividen cada 20 minutos si las condiciones les son favorables, por lo que una sola bacterias puede producir ingentes cantidades de descendientes en muy pocas horas. Se creyó durante mucho tiempo que sólo se reproducción asexualmente, pero hoy se conocen abundantes ejemplos se conjugación entre ellas, semejante a la de la reproducción sexual, al poder intercambiarse trozos de ADN, con lo que las bacterias resultantes tienen un material genético algo distinto. Se reproducen por bipartición simple, es decir, se parten en dos dividiendo equitativamente todo su contenido, incluido el ADN.
2011-0119 dios te bendiga
ResponderEliminarLa base del diagnóstico viral es la detección del virus o de sus componentes. El aislamiento del virus era la técnica standard de oro sobre la cual se medían todas las otras pruebas de diagnóstico viral, pero hoy en día con el desarrollo de las nuevas técnicas de Biología Molecular ya no es la más sensible. De igual forma el aislamiento de virus tiene una sensibilidad y una especificidad muy alta. Debido a que sólo se amplifica el virus, se aumenta la sensibilidad sin disminuir la especificidad. Sin embargo, existen algunas desventajas en el aislamiento del virus: El proceso suele ser lento, ya que demanda días a semanas para la identificación, y en consecuencia puede no estar disponible a tiempo para influir en la atención del paciente. Además, es un proceso laborioso y caro. Por otra parte requiere el uso de sistemas de cultivos adecuados, por ejemplo, se necesitan varias líneas celulares para la detección óptima de virus.
Los cultivos celulares son, pues, los biosubstratos más corrientemente empleados para la propagación de los virus.
Un cultivo celular es obtenido, de explantes de órganos o de embriones de animales.
Estas células obtenidas asépticamente se disocian tratándolas con una enzima (tripsina) que rompe el cemento intercelular. La suspención de celulas libres así obtenidas se coloca en la superficie plana de un recipiente de vidrio o plástico en donde las células se adhieren y multiplican formando una fina capa de células que se llama MONOCAPA CELULAR.
Esta monocapa de células crece en un medio de cultivo complejo que contiene albúmina, vitaminas, sales, glucosa, etc., en un sistema buffer. Se previene la contaminación bacteriana adicionándole al medio antibióticos adecuados.
Los cultivos celulares en monocapa son los más usados, aunque hay otros sistemas (cultivos en suspensión, explantos, cultivos de órganos, cultivos en microcarriers, etc.).
Los cultivos celulares se dividen en:
Cultivos Primarios: Se obtienen a partir de células, tejidos u órganos tomados directamente del organismo y pueden subcultivarse una o dos veces.
Líneas Celulares Diploides: Son aquellas que crecen en pasajes sucesivos hasta aproximadamente 50 subcultivos y que conservan por lo menos en un 75% el cariotipo correspondiente a la especie de que provienen.
Líneas Celulares Continuas: Permite un número finito de subcultivos y son heteroploides. Para considerar que se ha logrado establecer una línea continua, esta debe de haber sido subcultivada por lo menos 70 veces. Las líneas celulares continuas ofrecen las siguientes ventajas:
Disponibilidad para todos los investigadores de stocks de células idénticas, ya sea congeladas en ampollas o en monocapa de botella de cultivos, con la posibilidad de reconstituirlas cuando sea necesario.
Facilidad relativa del pasaje y mantenimiento en todos los laboratorios.
Libre de contaminación con agentes extraños.
2011-0119
ResponderEliminarCiclo reproductivo de los virus.
1-Fijación
2-Penetración
3-Desenvolvimiento
4-Síntesis (4a-Transcripción, 4b-Traducción, 4c-Replicación del genoma)
5-Ensamblaje
6-Liberación
El ciclo de replicación de los virus es el término utilizado para describir el ciclo de reproducción de los virus. Este consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden reproducirse por si mismos en el exterior; para ello requieren introducirse en células donde controlarán sus mecanismos reproductivos.
#1 Virologia
ResponderEliminarVirología es el estudio de virus y virus como agentes: clasificación y evolución, su estructura, sus formas de infectar y explotar las células para la reproducción del virus, las enfermedades que causan, las técnicas para aislar y la cultura de ellos y su uso en la investigación y terapia.
Virología es considerada una parte de la microbiología o de patología.
Una importante rama de la Virología es la clasificación de virus. Virus pueden clasificarse según la célula huésped que infectan:
virus animales, virus de plantas, hongos virus y bacteriófagos (virus que infectan bacterias, que son los virus más complejos).
Otra clasificación utiliza la forma geométrica de su cápside (a menudo una hélice o un icosaedro) o la estructura del virus (por ejemplo, la presencia o la ausencia de un envolvente de lípidos). Gama de virus en tamaño de unos 30 nm a aproximadamente 450 nm, lo que significa que la mayoría de ellos no puede verse con microscopios de luces.
El sistema de clasificación más útil y más ampliamente utilizado distingue virus segun el tipo de ácido nucleico que utilizan como material genético y el método de replicación viral que emplean coaxial células huésped en la producción de más virus:
• Virus de ADN (divididas en virus de ADN bicatenario y los virus de ADN monocatenario mucho menos común),
• Virus ARN (dividieron en virus de ARN monocatenario de sentido positivo, virus de ARN monocatenario de sentido negativo y los virus de ARN bicatenario mucho menos común),
• invertir transcribir virus (virus de ADN de transcripción inversa de bicatenario y monocatenario transcripción inversa RNA virus incluyendo retrovirus).
#2
EliminarEstructura General de los Virus
Un virus está compuesto de una molécula de ácido nucleico y una envoltura proteínica. Ésta es la estructura básica de un virus, aunque algunos de ellos pueden añadir a esto la presencia de alguna enzima, bien junto al ácido nucleico, como la transcriptasa inversa de los retrovirus, bien en la envoltura, para facilitar la apertura de una brecha en la membrana de la célula hospedadora.
A la unidad formada por el ácido nucleico y la envoltura proteínica se le denomina tambiénvirión.
El ácido nucleico es solamente de un tipo, ADN o ARN, nunca los dos.
Atendiendo al tipo de ácido nucleico se distinguen cuatro clases de virus:
- ADN de cadena doble
- ADN de cadena sencilla
- ARN de cadena doble
- ARN de cadena sencilla
La envoltura proteínica recibe el nombre de cápsida. Está formada por unas subunidades idénticas denominadas capsómeros. Los capsómeros son proteínas globulares que en ocasiones tienen una parte glicídica unida. Se ensamblan entre sí dando a la cubierta una forma geométrica.
Atendiendo la forma de la cápsida, se pueden distinguir los siguientes tipos de virus:
- Cilíndricos o helicoidales: los capsómeros, que son de un solo tipo, se ajustan entorno una hélice simple de ácido nucleico. Un ejemplo lo constituye el virus del mosaico del tabaco.
- Icosaédricos: los capsómeros, que suelen ser de varios tipos, se ajustan formando un icosaedro regular (es decir, 20 caras triangulares y 12 vértices), y dejando un hueco central donde se sitúa el ácido nucleico fuertemente apelotonado. Algunos forman poliedros con más caras que el icosaedro, y algunos presentan fibras proteicas que sobresalen de la cápsida.
Un ejemplo lo constituyen los adenovirus, entre los que se encuentran los virus de los resfriados y faringitis.
- Complejos: con pequeñas variantes, responden a la siguiente estructura general.
- Una cabeza de estructura icosaédrica que alberga el ácido nucleico.
- Una cola de estructura helicoidal que constituye un cilindro hueco.
- Un collar de capsómeros entre la cabeza y la cola.
- Una placa basal, al final de la cola, con unos puntos de anclaje que sirven para fijar el virus a la membrana celular.
De la placa salen también unas fibras proteicas que ayudan a la fijación del virus sobre la célula hospedadora.
Como ejemplo de este tipo de virus se encuentran la mayor parte de los virus bacteriófagos (que infectan las bacterias).
#3
EliminarSimetrías Virales
La simetría es la forma que adopta un virus en el espacio y está dad por la estructura de su nucleocápside. La simetría puede ser helicoidal, icosaédrica o compleja.
Los virus con simetría helicoidal presentan una nucleocápside cilindrica que puede estar extendida como en el virus del mosaico del tabaco. Este es un virus desnudo.
En otros casos la nucleocápside está arrollada sobre sí misma y recubierta por una envoltura, como en el virusInfluenza.
Los virus con simetría icosaédrica son poliedros regulares con 20 caras triangulares, 30 aristas y 12 vértices. Son ejemplos de virus con simetría icosaédrica desnudos el virus de la poliomielitis o losadenovirus. Los herpesvirus y togavirus son virus con simetría icosaédrica y recubiertos por una envoltura.
Se denominan virus de simetría compleja a aquellos que presentan una nucleocápside helicoidal o icosaédrica recubierta por una envoltura laxa. Ejemplos de éstos virus son los poxvirus (forma de ladrillo), los rhabdovirus (forma de bala) y algunos bacteriófagos que poseen una cabeza con simetría icosaédrica y una cola con simetría helicoidal.
#4
EliminarTamaño de los Virus
Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus.
Su pequeño tamaño explica lo tardío del descubrimiento de estos agentes. La primera referencia sobre la existencia de los virus se debe al botánico ruso Dimitri Ivanovski en 1892. Este investigador buscaba el agente causante de la enfermedad denominada mosaico del tabaco, y llegó a la conclusión de que debía tratarse de una toxina o de un organismo más pequeño que las bacterias, pues el agente atravesaba los filtros que retenían las bacterias. Denominó a estos agentes patógenos virus filtrables.
En 1897, el microbiólogo holandés Martinus Beijerink realizó experimentos similares a los de Ivanovski, y llegó a desechar la idea de las toxinas, pues se trataba de un agente capaz de reproducirse, ya que mantenía su poder infeccioso de unas plantas a otras, sin diluirse su poder patógeno. Poco después, los microbiólogos alemanes Frederick Loeffler y Paul Frosch descubrieron que la fiebre aftosa del ganado era producida por un virus filtrable que actuaba como un agente infeccioso.
En la década de los 30, con el uso de filtros de tamaño de poro inferior, con las técnicas de cultivo celular in vitro que permitían la obtención de gran cantidad de virus, con la ultracentrifugación y finalmente con el microscopio electrónico y la difracción de rayos X, se logró visualizar a estos agentes.
Observaciones:
En la tabla correspondiente a esta diapositiva, disponemos un cuadro correspondiente al tamaño de cada uno de los cuales corresponden a esta categoría.
# 5
EliminarFamilia de Virus
Imagen:
Nos muestras las diferentes familiar relacionadas a los virus las cuales conoceremos a continuación entre ellas están:
1- Picornaviridae: es una familia de virus infectivos para animales. Contienen un genoma ARN monocatenario positivo y por lo tanto se incluyen en el Grupo IV de la Clasificación de Baltimore. El genoma ARN es inusual porque tiene una proteína en el terminal 5' que se utiliza como iniciador de la transcripción por la ARN polimerasa. El nombre se deriva de "pico", que significa pequeño, con lo que "picornavirus" significa literalmente "virus RNA pequeños".
2- Caliciviridae: es una familia de virus infectivos para animales y causantes de gastroenteritis en humanos. Los calicivirus han sido encontrados en la mayoría de los animales domésticos y muchos silvestres, como cerdos, conejos, gallinas y anfibios. Contienen un genoma ARN monocatenario positivo y por lo tanto se incluyen en el Grupo IV de la Clasificación de Baltimore.
3- Hepadnaviridae es una familia de virus que causan infecciones en hígado de humanos y de animales. Los hepadnavirus tienen un genoma muy corto de ADN parcialmente de doble hélice y parcialmente de hélice simple, circular. Como su replicación involucra un ARN intermedio, se incluyen en el Grupo VII de la Clasificación de Baltimore.
4- Reoviridae es una familia de virus que pueden afectar el sistema gastrointestinal (tales como rotavirus ) y tracto respiratorio . Los virus de la familia Reoviridae han genomas que consiste en segmentar, ARN de doble cadena (dsRNA). El nombre "Reoviridae" se deriva de las vías respiratorias entéricas virus huérfanos.
5- Rhabdoviridae es una familia de virus infectivos para animales y plantas. Entre los animales infectados están insectos, peces y mamíferos, incluidos los humanos. Estos virus presentan una forma alargada parecida a una bala. Poseen un genoma de ARN monocatenario de sentido negativo por lo que se incluye en el Grupo V de la Clasificación de Baltimore.
6- El Togaviridae es una familia de virus que incluye los siguientes géneros:
Género Alphavirus; especies típicas: Sindbis virus, Virus de la encefalitis equina oriental, Virus de la encefalitis equina occidental, Virus de la encefalitis equina venezolana, Virus del Río Ross, Virus O'nyong'nyong.
Género Rubivirus; especies típicas: Rubéola virus.
7- Los Coronavirus son un género de virus ARN de vertebrados de Coronaviridae.1 Son virus envueltos con un genoma de ARN de cadena sencilla con polaridad positiva y simetría helicoidal. El tamaño genómico de los coronavirus varía entre 16 a 31 kilobases, un virus de extraordinaria longitud.
8- Bunyavirus, género de virus ARN de vertebrados de la familia Bunyaviridae.
9- Papovaviridae es una familia de virus pequeños (apróx. 50 nm) infectivos para una gran variedad de animales, incluyendo los humanos. Su nombre implica las patología que causa: papilomavirus + poliomavirus + agente vacuolante. Tienen una amplia distribución mundial.
10- Los paramixovirus son miembros de la familia de virus paramyxoviridae, que poseen un genoma con ARN de cadena sencilla de polaridad negativa como ácido nucleico; albergan dicha información genética en una cápside cubierta de envoltura viral y estructuralmente definida por una simetría helicoidal, de un tamaño de 125 a 250 x 18 nm, con envoltura, pleomórfico, y un filamentoso nucleocápside; y por ensamblar los viriones maduros en el citoplasma como compartimento celular.
11- Los Orthomyxoviridae son una familia de virus RNA que infectan a los vertebrados. Incluyen a los virus causantes de la gripe.
Eliminar12- Un retrovirus es un virus de ARN que se replica en una célula huésped a través del proceso de transcripción inversa . En primer lugar, utiliza su propio transcriptasa inversa enzima para producir ADN de su ARN del genoma, la inversa de la pauta habitual, por lo tanto retro (hacia atrás).
13- Flavivirus : es un género de virus ARN pertenecientes a la familia Flaviviridae. Los Flavivirus son virus con envoltura, la simetría de la nucleocápside icosaedrica, y cuyo material genético reside en una única cadena de ARN de polaridad positiva.
14- Parvovirus, a veces truncada a "parvo", es a la vez el nombre común en Inglés casualmente se aplica a todos los virus de la Parvoviridae familia taxonómica y también el nombre taxonómico del parvovirus género dentro de la Parvoviridae familia.
15- Los adenovirus (Adenoviridae) son una familia de virus que infectan tanto humanos como animales. Son virus no encapsulados de ADN bicatenario que pueden provocar infecciones en las vías respiratorias, conjuntivitis, cistitis hemorrágica y gastroenteritis. Los adenovirus también se utilizan para obtener ADN para la terapia génica.
16- El Arenavirus es un género de virus de la familia Arenaviridae.La especie tipo es el virus de la coriomeningitis linfocítica (LCMV); es también de las spp. responsables de la fiebre Lassa. Arena, viene del latín con la familia de términos del árido.
17- Los herpesvirus o virus de la familia Herpesviridae, deben su nombre al término griego herpein (ἕρπειν), reptar o arrastrar, haciendo alusión a la facultad de estos agentes infecciosos microscópicos de ser fácilmente contagiados y transmitidos de una persona a la otra y de recurrencia crónica. Los herpesvirus forman una familia de virus divididos en tres subfamilias: alpha, beta y gammaherpesvirinae, ubicados con base en la arquitectura del virión y ciertas propiedades biológicas comunes.
18- Poxviridae es una familia de virus de ADN relacionados entre sí llamados poxvirus, infectivos para animales vertebrados (8 de los 11 géneros, por ejemplo, poxvirus de la gallina) y a los invertebrados (3 géneros). Tienen un genoma ADN bicatenario y por lo tanto pertenece al Grupo I de la Clasificación de Baltimore.
19- Filoviridae es una familia de virus del orden Mononegavirales. Se caracterizan por poseer como genoma RNA monocatenario de sentido negativo, y por infectar a primates. Agrupa, entre otros, al virus Ébola y al virus Marburg.
#6
EliminarDiagnostico de la Virosis
Los virus son organismos tan pequeños, que su tamaño les permite pasar a través de los filtros que retienen bacterias. Las características básicas para identificar a un virus son: la composición del ácido nucleico, la disposición de la cápside, la presencia o ausencia de una envoltura, el tamaño del virus y el huésped.
MUESTRAS PARA DIAGNOSTICO:
Las muestras deben tomarse precozmente en la fase aguda de la infección. Deben transladarse al laboratorio a la mayor brevedad posible, en medio de transporte. Las muestras más comunes son:
• Torundas nasofaríngeas: adecuados para el diagnóstico de enterovirus, adenovirus, y herpes simplex. Las nasofaringeas se prefieren para el virus respiratorio sincicial y la mayoria de los otros respiratorios. Las nasales son las mejores para los rinovirus.
• Aspirados faríngeos: son mejores que los hisopos (torundas) nasofaringeos, pero estos últimos son mas prácticos.
• Torundas rectales y muestras de heces: Aquí se muestran virus no cultivables como rotavirus, o adenovirus, que se detectan por microscopía electrónica o con técnicas de detección de antígenos.
• Secreciones, Sangre o Líquido cefalorraquídeo.
• En algunas virosis excepcionalmente el diagnóstico solo puede efectuarse en Biopsia de cerebro, como en la encefalitis por herpes simplex; en Huéspedes inmunodeprimidos y con citomegalovirus, el virus solo puede identificarse en el órgano afectado.
DIAGNOSTICO VIRAL
Observación del efecto citopático, en cultivo de tejidos inoculados con ciertos virus. La etiología de un determinado síndrome viral puede demostrarse recuperando el agente causal en tejidos celulares de diversos tipos (por ej: riñón de mono, pulmón humano fetal, amnios humano o células tumorales humanas). Los huevos embrionados actualmente solo se utilizan para cultivar el virus cuando se desea preparar vacunas. El empleo de animales de experimentación es excepcional en el diagnóstico.
• Las células primarias de riñón de mono son excelentes para cultivar mixovirus, paramixovirus y muchos enterovirus; también para algunos adenovirus.
• Las células diploides fetales humanas son células fibroblásticas en las que crece un amplio espectro de virus, como el de herpes simple, varicela zoster, adenovirus, picornavirus, y sólo en éstas pueden cultivarse los citomegalovirus.
• Las células HEP-2 son epiteliales provenientes de un cancer humano, útiles para recuperar el virus respiratorio sincitial, adenovirus, y herpes simple.
• Células mononucleares normales normales humanas de sangre periférica, que no suelen estar disponibles, permiten el cultivo del virus ( SIDA ) de la inmunodeficiencia, el coxsackie A y los togavirus.
El crecimiento de un virus se detecta mediante la observación de cambios en la monocapa del cultivo celular, denominado efecto citopático (ECP). Cuando aparecen cambios en la morfología celular, lisis celular, vacuolización, formación de cincicios y existen cuerpos de inclusión, se produce un efecto citopático característico. Los cuerpos de inclusión son cambios histológicos que se producen en las células por los componentes virales o por variaciones en las estructuras celulares.
EliminarUn técnico experto puede distinguir los efectos citopáticos característicos de las principales familias de virus. Según el cultivo celular en el que aparece el ECP y el tipo de crecimiento viral, se puede realizar un diagnóstico de presunción de un gran número de virus de importancia clínica.
Algunos virus no producen rápidamente ECP en las líneas celulares que se suelen utilizar en los laboratorios de virología clínica. No obstante se pueden determinar por otras técnicas, como la interferencia en la replicación de otros virus (por ej los picornavirus no pueden replicarse en células previamente infectados con el virus de la rubeola, lo que se denomina interferencia heteróloga).
1.2. Fijación de eritrocitos en cultivos celulares infectados con virus. En el caso de virus que dan lugar a este fenómeno, como mixovirus o virus de la parotiditis, se observa una hemaglutinación al adicionar eritrocitos a la monocapa de células.
1.3. Detección de antígenos virales. Cuando los virus se pueden detectar antes de desarrollar el efecto citopático, se utilizan anticuerpos monoclonales dirigidos contra antígenos tempranos y técnicas de inmunofluorescencia o inmunoperoxidasa.
1.4. Inmunofluorescencia directa, utilizando células del paciente, como leucocitos, orina o mucosa oral o genital, para buscar virus.
1.5. Detección de ácidos nucleicos virales por reacción de polimerasa en cadena, las cuales permiten amplificar una porción pequeña de ácido nucleico en un período de tiempo corto.
1.6. Microscopía electrónica. Cuando algunos virus comunes no se aislan con los métodos de cultivo habituales, ni pueden diagnosticarse por técnicas serológicas, pueden estudiarse con otras técnicas, como la microscopía electrónica. Tal es el caso de los calcivirus, los minirrotavirus, los astrovirus y los parvovirus del cerebro. Sin embargo, hay tres grandes dificultades para su uso: el instrumento es caro y es difícil su mantenimiento. Por otro lado, deben estar presentes grandes cantidades de particulas virales en la muestra para poder detectarse. Finalmente, la morfología no es suficientemente diferencial para un diagnóstico preciso.
#7
EliminarEtapas de la Replicacion Viral
Ciclo reproductivo de los virus.
1-Fijación
2-Penetración
3-Desenvolvimiento
4-Síntesis (4a-Transcripción, 4b-Traducción, 4c-Replicación del genoma)
5-Ensamblaje
6-Liberación
El ciclo de replicación de los virus es el término utilizado para describir el ciclo de reproducción de los virus. Este consta generalmente de las siguientes fases: fijación y entrada en la célula, eclipse, multiplicación y liberación del virus. Los virus son formas acelulares que no pueden reproducirse por si mismos en el exterior; para ello requieren introducirse en células donde controlarán sus mecanismos reproductivos.
#8
EliminarAdhesión
La adhesión es la propiedad de la materia por la cual se unen y plasman dos superficies de sustancias iguales o diferentes cuando entran en contacto, y se mantienen juntas por fuerzas intermoleculares.
Mecanismos de adhesion
La cohesión es la causa de que el agua forme gotas, la tensión superficial hace que se mantengan esférica y la adhesión la mantiene en su sitio.
Las gotas de agua son más planas sobre la flor de Hibiscus ya que tienen mejor adhesión.
Cinco mecanismos han sido propuestos para explicar por qué un material se adhiere a otro.
Adhesión mecánica
Los materiales adhesivos rellenan los huecos o poros de las superficies manteniendo las superficies unidas por enclavamiento. Existen formas a gran escala de costura, otras veces a media escala como el velcro y algunos adhesivos textiles que funcionan a escalas pequeñas. Es un método similar a la tensión superficial.
Adhesión química
Dos materiales pueden formar un compuesto químico al unirse. Las uniones más fuertes se producen entre átomos donde hay permutación (enlace iónico) o se comparten electrones (enlace covalente). Un enlace más débil se produce cuando un átomo de hidrógeno que ya forma parte de una partícula se ve atraída por otra de nitrógeno, oxígeno o flúor, en ese caso hablaríamos de un puente de hidrógeno.
Adhesión dispersiva
En la adhesión dispersiva, dos materiales se mantienen unidos por las fuerzas de van der Waals: la atracción entre dos moléculas, cada una de las cuales tiene regiones de carga positiva y negativa. En este caso, cada molécula tiene una región de mayor carga positiva o negativa que se une a la siguiente de carga contraria. Este efecto puede ser una propiedad permanente o temporal debido al movimiento continuo de los electrones en una región.
En la ciencia de superficies el término "adhesión" siempre se refiere a una adhesión dispersiva. En un sistema sólido-líquido-gas normal (como una gota de un líquido sobre una superficie rodeada de aire) el ángulo de contacto es usado para cuantificar la adhesividad. En los casos donde el ángulo de contacto es bajo la adhesión está muy presente.
Esto se debe a que una mayor superficie entre el líquido y el sólido conlleva una energía superficial mayor.
Adhesión electrostática
Algunos materiales conductores dejan pasar electrones formando una diferencia de potencial al unirse. Esto da como resultado una estructura similar a un condensador y crea una fuerza electrostática atractiva entre materiales. mi con diversos materiales les pueden ayudar.
#9
EliminarPenetración
La penetración es el acto de introducir un elemento dentro de otro elemento.
Imagen:
Muestras los diferentes procesos que experimenta la penetración en estos procesos correspondientes a los virus.
#10
EliminarPenetración de los Bacteriófagos
Los virus son moléculas de DNA o RNA rodeadas por una envoltura proteica que necesitan células viables para poder replicarse. Los virus utilizan la maquinaria metabólica de las células para sintetizar su material genético y proteínas de la envoltura. Existen distintos tipos de virus que pueden infectar células procariontes o células eucariontes. Los bacteriófagos o fagos son virus que se reproducen en células procariontes.
El genoma de los fagos puede ser RNA simple cadena (MS2, Qß), RNA doble cadena (phi 6), DNA simple cadena (phi X174, fd, M13) o DNA doble cadena (T3, T7, lambda , T5, Mu, T2, T4). Estos ácidos nucleicos pueden contener bases inusuales que son sintetizadas por proteínas del fago. En los T-pares el genoma no contiene citosina sino 5'-
hidroximetilcitosina, mientras que en otros tipos de fago alguna de las bases esta parcialmente sustituida.
#11
EliminarReplicacion y Maduracion
Replicacion:
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante.
Maduración y ensamblaje
Cuando se han sintetizado un número crítico de varios componentes virales, éstos se ensamblan en partículas víricas maduras en el núcleo y/o citoplasma de la célula infectada. En este ensamblaje y al contrario de lo que ocurre en los bacteriófagos, no están implicados enzimas biosintéticos especiales, sino que ocurre de forma espontánea debido a la gran afinidad de la cápsida con el ácido nucleico viral. La maduración se puede definir como la fase de la infección viral durante la cual los componentes estructurales del virión se producen y ensamblan con el nuevo ácido nucleico viral para formar la nucleocápsida.
#12
EliminarMaduración y Salida de Virus Desnudos y Envueltos
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. (Figura 1).
(B) Entada vía endosomas en la superficie celular (figura 2)
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Virus no envueltos o desnudos
Los virus no envueltos o desnudos pueden cruzar la membrana plasmática directamente o pueden ser tomados en endosomas. Si son transportados en endosomas, luego cruzan (o destruyen) la membrana de dichas estructuras.
#13
EliminarSalida de un Virus Envuelto
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
#14
EliminarAntivirales
Los antivirales son un tipo de medicamento usado para el tratamiento de infecciones producidas por virus. Tal como los antibióticos (específicos para bacteria), existen antivirales específicos para distintos tipos de virus. No sin excepciones, son relativamente inocuos para el huésped, por lo que su aplicación es relativamente segura. Deben distinguirse de los viricidas, que son compuestos químicos que destruyen las partículas virales presentes en el medio ambiente.
El primer antiviral exitoso, el aciclovir, es un análogo a los nucleósidos y es efectivo contra el herpesvirus. La primera sustancia antiviral en ser aprobada para tratar el VIH, la zidovudina (AZT), es también un nucleósido análogo.
Un mejor conocimiento de la acción de la transcriptasa inversa ha conducido a mejores análogos a nucleósidos para tratar las infecciones del VIH. Uno de estos medicamentos, la lamivudina, ha sido aprobado para tratar la hepatitis B, que utiliza la transcriptasa inversa como parte de su proceso de replicación. Los investigadores han ido más lejos y han desarrollado inhibidores que no parecen nucleósidos, pero pueden seguir bloqueando la transcriptasa inversa.
Otros objetivos a ser considerados por los antivirales VIH incluyen la RNasa H, que es un componente de la transcriptasa inversa que rompe el ADN sintetizado del ARN viral original y la integrasa, que empalma el ADN sintetizado con el genoma de la célula huésped.
#15
EliminarMecanismo de Acción de los Retrovirales
Los retrovirus son pequeños microorganismos que contienen una única cadena de ARN. Su ciclo vital se basa en la conversión de su ARN geonómico en doble cadena de ADN y posteriormente en la integración de éste último en los cromosomas de la célula huésped en lo que se denomina provirus endógeno. De forma ocasional algunas secuencias celulares son recombinadas con el provirus y a su vez estas recombinaciones pueden transferirse a otra población celular infectada, lo que puede tener importantes consecuencias en las funciones celulares.
Peyton Rous identificó el primer retrovirus en 1911 cuando se encontró un agente transmisible capaz de causar un tipo de sarcoma en pollos. Fue tras el descubrimiento del enzima transcriptasa inversa por Temin cuando se pudo ahondar en sus mecanismos de acción y en su manipulación experimental.
Como se comentó al inicio de esta exposición fue hacia 1976 cuando se demostró que los supuestos oncogenes que portaban algunos de estos virus tenían una contrapartida en humanos como genes normales a los que se denominaron protooncogenes.
Otro elemento esencial en el estudio de los retrovirus lo constituyó el descubrimiento del primer retrovirus infeccioso humano el HTLV-1 al que luego se le sumaría el HIV.
La familia de los retrovirus ha sido subclasificada en tres subfamilias: Oncornavirus (que incluye al virus del tumor mamario murino, el virus de la reticuloendoteliosis aviar, virus de la leucemia humana de células T), Lentivirus (que incluye los virus de las inmunodeficiencias) y los Spumavirus.
#16
EliminarRetrovirales
Retroviridae es una familia de virus que comprende los retrovirus. Son virus con envoltura que presentan un genoma de ARN monocatenario de polaridad + y se replican de manera inusual a través de una forma intermedia de ADN bicatenario.
Este proceso se lleva a cabo mediante una enzima: la retrotranscriptasa o transcriptasa inversa, que dirige la síntesis de ADN a través de ARN y posee una importancia extraordinaria en la manipulación genética. Una vez se ha pasado de ARN monocatenario a ADN se inserta dentro del ADN propio de la célula infectada donde se comporta como un gen más (véase Ciclo reproductivo de los virus). Por tanto, se incluyen en el Grupo VI de la Clasificación de Baltimore.
Los retrovirus son responsables de muchas enfermedades, incluyendo algunos cánceres y el sida (VIH). Existen diversos grupos de investigación que han intentado modificar genéticamente los retrovirus para usarlos en terapia génica como vectores, pero se han encontrado con diversos problemas.
#17
EliminarBiones Acelulares
Virus: es un agente infeccioso microscópico que sólo puede multiplicarse dentro de las células de otros organismos.Los virus infectan todos los tipos de organismos, desde animales y plantas, hasta bacterias y arqueas.
Los virus son demasiado pequeños para poder ser observados con la ayuda de un microscopio óptico, por lo que se dice que son submicroscópicos; auque existen excepciones entre los Virus nucleocitoplasmáticos de ADN de gran tamaño, tales como el Megavirus chilensis, el cual se logra ver a traves de microscopía óptica.
Un virusoide o ARN satélite es una pequeña molécula patógena de ARN (ácido ribonucleico) que se encuentra encapsulada en ciertos virus de plantas con secuencias de ARN de mayor tamaño, lo que se denomina un virus asistente, el cual hospeda al virusoide y es necesario para que éste produzca una infección exitosa.
Los viroides son agentes infecciosos que, al igual que los virus, tienen un ciclo extracelular que se caracteriza por la inactividad metabólica y un ciclo intracelular en el que causan infección al huésped susceptible, pero que a diferencia de los virus, los viroides no poseen proteínas ni lípidos y están constituidos por una cadena cíclica corta de ARN, circular o con forma de varilla, (que no codifica proteínas). Es importante decir que tanto su forma intracelular como extracelular son las mismas (ARN desnudo), los mecanismos por los cuales éstos logran causar infección están relacionados con la autocatálisis de su material genético.
Los priones son los agentes causantes de un grupo de patologías neurodegenerativas letales características de mamíferos, también conocidas como encefalopatías espongiformes transmisibles. Estos agentes son capaces de propagarse dentro de un mismo huesped causando una lesión espongiótica y de transmitirse de huesped a huesped con elevados tiempos de incubación.
Transposon: Secuencia de DNA que puede moverse de un lugar a otro del cromosoma, insertar copias adicionales de ella misma en otros puntos o pasar de un cromosoma a otro. Tramo de DNA que puede incorporarse en otras moléculas de DNA en lugares donde no hay homología de secuencia. Tienen un tamaño entre 1 y 40 kb. Codifican todas las enzimas necesarias para su inserción. Pueden desplazarse dentro de un cromosoma o entre cromosomas.
Los plásmidos son moléculas de ADN extracromosómico circular o lineal que se replican y transcriben independientes del ADN cromosómico. Están presentes normalmente en bacterias, y en algunas ocasiones en organismos eucariotas como las levaduras. Su tamaño varía desde 1 a 250 kb.
#18
EliminarViroides 1
Los viroides, al igual que los virus son parásitos intracelulares estrictos, es decir, dependen de la célula que infectan para su multiplicación, pero aquí acaban las similitudes. El virus más pequeño tiene un genoma de unas 1500 bases. En cambio el genoma de un viroide está comprendidos entre 250 y 400 nucleótidos (unas 20 veces más pequeños). El genoma de los virus puede ser de ADN o de ARN de doble hebra o de cadena simple, y a pesar que parasitan la maquinaria celular del huésped, siempre codifican alguna proteína. Los viroides en cambio siempre son estructuras de ARN de una sola hebra, circulares y su secuencia no codifica ninguna proteína. Los virus tienen una cápsida o cobertura, los viroides en cambio son la hebra de ARN desnuda.
Los viroides fueron descubiertos en los años 1970 por Theodore Otto Diener, al demostrar que el agente de la enfermedad del tubérculo fusiforme de la patata era una entidad más pequeña que los virus. La comunidad científica se mostró reacia al descubrimiento y en un principio se achacó a que pudiera ser un ARN satélite (secuencias de ARN que parasitan a otros virus… si, algunos virus también tienen sus propios parásitos). Pero no era el caso. Y quizás esto explica por que se habla tan poco de ellos en los libros de biología. Solo existen en plantas. No existen viroides en animales, aunque el virus de la hepatits delta tiene ciertas similitudes con los viroides, no se ha podido demostrar un origen evolutivo común.
#19
EliminarViroides II
Los viroides son los agentes infecciosos de menor complejidad genética y estructural conocidos y representan una forma extrema de parasitismo. Están constituidos exclusivamente por moléculas de ARN de cadena simple, cerradas covalentemente o con forma de bastón de bajo peso molecular (246 a 1994 nucleótidos). Carecen de actividad de ARN mensajero y se replican de forma autónoma, utilizando el sistema de transcripción de la célula suceptible.2 Los viroides se encuentran, casi exclusivamente, en el núcleo de las células infectadas; se desconoce el modo en que se replican pero se sabe que el ARN que los constituyen no funciona como ARN mensajero y tampoco se traduce a enzimas que participen en su propia replicación. Los viroides no codifican para ninguna proteína y llevan a cabo su replicación en la célula hospedadora a través de la RNA polimerasa II codificada por la célula hospedadora aunque también se ha propuesto la posibilidad de que lo haga gracias a una RNA polimerasa dependiente de RNA presente en algunas células eucariotas (Cann, 2001). Dada su localización, se presume que causan la enfermedad interfiriendo con la regulación génica de la célula hospedadora en la etapa de corrección del ARN mensajero celular: particularmente en la eliminación de intrones y de empalmes de exones, presentando secuencias muy similares a los intrones que normalmente son cortados. Sus ARN se replican varias veces.
#20
EliminarOtros biones Acelullares
Un virus satélite es un agente subvírico formado por ácido nucleico y que depende de la coinfección de la célula huésped por un virus (helper o máster) para reproducirse; obteniendo de ellos lasenzimas faltantes en su genoma, las cuales son necesarias para lograr replicarse.
Un virusoide o ARN satélite es una pequeña molécula patógena de ARN (ácido ribonucleico) que se encuentra encapsulada en ciertos virus de plantas con secuencias de ARN de mayor tamaño, lo que se denomina un virus asistente, el cual hospeda al virusoide y es necesario para que éste produzca una infección exitosa.
La McIntosh Roja o Mac es un tipo de manzana de piel roja y verde. Se recoge a finales de septiembre y es el cultivo más popular de Nueva Inglaterra, muy conocida por la compota rosada que se hace a partir de manzanas McIntosh sin pelar.
Cada manzana de McIntosh tiene un linaje directo a un solo árbol descubierto en 1796 por John McIntosh en su granja en Dundela, una aldea situada en el condado de Dundas en la provincia canadiense de Ontario, cerca de Prescott.
Los plásmidos son fragmentos extracromosómicos de ácidos nucléicos (ADN o ARN) que aparecen en el citoplasma de algunos procariotas. Son de tamaño variable aunque menor que el cromosoma principal.
#21
EliminarPriones I
Los priones son los agentes causantes de un grupo de patologías neurodegenerativas letales características de mamíferos, también conocidas como encefalopatías espongiformes transmisibles. Estos agentes son capaces de propagarse dentro de un mismo huesped causando una lesión espongiótica y de transmitirse de huesped a huesped con elevados tiempos de incubación. A diferencia de virus y viroides, son resistentes a tratamientos inactivantes de ácidos nucléicos, pero comparten con éstos la existencia de una variabilidad de inóculos dentro de la misma especie (diferenciables por el patrón de la lesión y la magnitud del tiempo de incubación) y de una infectividad sujeta a barrera de especie (Chandler, 1961; Alper y cols., 1967; Hunter, 1972; Prusiner, 1982; Bruce y Fraser, 1991; Bessen y Marsh, 1992).
La búsqueda de la entidad molecular constitutiva de este agente reveló como componente mayoritario, si no único, una proteína: PrPSc, proteína del prion de scrapie),y la ausencia de un ácido nucleico específico (Prusiner 1982, 1991). Con estas premisas estructurales unidas a la capacidad de infección, Prusiner acuña el término prion (particula infecciosa de naturaleza proteica) para diferenciarlo de virus y viroides. Dadas las características poco convencionales de los priones se han elaborado numerosas hipótesis sobre su estructura (Dickinson y Outram, 1988; Prusiner, 1991; Weissmann, 1991). En la actualidad la hipótesis con mayor grado de aceptación es la conocida como "sólo proteína", delineada inicialmente por Griffith (1967) y, formalmente enunciada y actualizada por Prusiner (1991, 1997). Tras la descripción de proteínas de diferente secuencia pero similar comportamiento en levadura así como los avances interdisciplinares realizados en el conocimento de estos agentes en la última década, el concepto de prion ha sido acotado adquiriendo una defición más precisa y generalizable (Wicker y Masison, 1996; Lindquist S., 1997; Prusiner, 1997). Así, se denomina prion a la forma alterada de una proteína celular funcional (PrP en mamíferos) que ha podido perder su función normal pero que ha adquirido la capacidad de transformar la forma normal en patológica.
#22
EliminarPriones II
Han sido definidos como pequeñas partículas proteináceas que son resistentes a la inactivación por procedimientos que modifican los ácidos nucléicos.
Otra definición de prion es la forma alterada de una proteína celular funcional que ha podido perder su función normal pero que ha adquirido la capacidad de transformar la forma normal en patológica.
Los mamíferos tienen en su genoma genes que codifican la “proteína priónica”(PrP), y esta proteína en condiciones normales tiene una configuración espacial de tipo hélices alfa, que es susceptible a la lisis por proteasas y no es patógena. Los priones no son seres vivos ni tienen la capacidad de infección como tal.
La patogénesis de estas proteínas se produce cuando tienen bien una mutación (esporádica o adquirida genéticamente) o bien un cambio conformacional inducido por PrP-Sc (patógena) de otro individuo.
El cambio conformacional o las mutaciones hacen que la PrP tenga una estructura tridimensional diferente constituida por hélices beta con agregados amorfos insolubles (PrP-Sc), de manera que se hace imposible su lisis por las proteasas, produciéndose su acúmulo en los lisosomas de las células de los tejidos afectados y originando así las enfermedades priónicas.
Hugo Avila Lamarche(2011-0232)(Grupo No.3) La virología (rama de la microbiologia) es el estudio de los virus: su estructura, clasificación y evolución, su manera de infectar y aprovecharse de las células huésped para la reproducción del virus, su interacción con los organismos huéspedes, su inmunidad, la enfermedad que causan, las técnicas para su aislamiento, cultivo y su uso en investigación y terapia. La virología es considerada un sub-campo de la microbiología y la medicina. La virologia también es una rama taxonómica de la biología para estudiar los virus y todos sus componentes.
ResponderEliminarSu investigación incluye:
* la replicación viral
* los patógenos virales.
* la inmunología viral.
* las vacunas virales.
* los métodos de diagnóstico.
* la quimioterapia antiviral.
* las medidas de control de una infección.
* los diferentes signos que manifiestan la presencia de virus.
yuleika veloz 2011-0481
ResponderEliminarLas células eucariotas surgieron de antepasados procariotas. El origen de los eucariotas se encuentra en sucesivos procesos simbiogenéticos (procesos simbióticos que culminan en la unión de sus simbiontes, estableciéndose una nueva individualidad de los integra) entre diferentes bacterias.
La teoría endosimbiótica postula que algunos orgánulos propios de las células eucariotas, especialmente plastos y mitocondrias, habrían tenido su origen en organismos procariotas que después de ser englobados por otro microorganismo habrían establecido una relación endosimbiótica con éste. Se especula con que las mitocondrias provendrían de protebacterias alfa (por ejemplo, rickettsias) y los plastos de cianobacterias.
Mas tarde con el tiempo y por evolución celular estas bacterias quedarían transformadas en Mitocondrias (bacterias aerobias) y en Cloroplastos (bacterias Fotótrofas), no tiene que ver con la evolución de las algas sino con la aparición de Mitocondrias y cloroplastos, los 2 organelos membranosos que se encuentran únicamente en células eucariotas poseen ADN de tipo procariota, ARN y Ribosomas dentro del mismo, pero la Teoría Endosimbiótica es mas aplicada en las Mitocondrias porque aparte de poseer 1sola molécula de ADN de tipo Procariota, ARN y Ribosomas en el lado interno de las "Crestas Mitocondriales que son proyecciones incompletas hacia la Matriz mitocondrial de la membrana interna" esta la Cadena Oxidativa o respiratoria.
Estas se diferencian principalmente, en que mayoría de las células eucariotas se distinguen de las procariotas
por la existencia, en su citoplasma, de varios millares de estructuras organizadas, los orgánulos, de cuyo tamaño viene a ser el de una célula procariota.
Célula Bacteriana:
Las células bacterianas esta constituida principalmente, por elementos facultativos y obligados.
Elementos obligatorios : están presentes en todas las bacterias y son indispensables para la vida de la propia bacteria estos son los siguientes:
- Pared celular
- Membrana citoplasmática
- Citoplasma
- Ribosomas
- Región nuclear
Elementos facultativos: pueden estar o no presentes en la bacteria y son:
- Cápsula
- Flagelos
- Fimbrias o Pili(pelo)
- Endosporas
- Inclusiones citoplasmáticas
yuleika veloz 2011-0481
ResponderEliminarLa pared gram positiva y gram negativa
La gram positiva mide de unos 15 a 20 nanometros de diametro y la gram negativa mide 2 nanometros de diametro.
La pared gram positiva, poseen una pared celular interna y una pared de peptidoglucano(es un exoesqueleto que da consistencia y forma esencial para replicación y supervivencia de la bacteria), no tiene membrana externa, no tiene espacio periplasmático, la red de mureína está muy desarrollada y llega a tener hasta 40 capas, conservan el complejo yodocolorante.
La pared gram negativa, poseen una pared celular más compleja:
pared celular interna, pared de peptidoglucano, bicapa lipídica externa, tiene membrana externa que forma un saco rígido alrededor de la bacteria, mantiene estructura y es barrera impermeable a macromoléculas, ofrece protección en condiciones adversas, tiene espacio periplasmático, pueden ser anaerobios o aerobios y poseen proteínas con concentraciones elevadas.
Coloración del Gram
La tinción de Gram o coloración de Gram es un tipo de tinción diferencial empleado en Bacteriología para la visualización de bacterias, sobre todo en es una prueba potente y rápida que nos permite diferenciar dos clases de bacterias.
Es una prueba potente y rápida que nos permite diferenciar dos clases de bacterias estas son: Bacterias gram positivas y las gram negativas.
Las grampositivas se tiñen de morado ya que el colorante se queda atrapad en la capa gruesa de peptidoglucanos que rodea a la célula
Las gramnegativas tienen una capa de petidoglucano mucho más delgada es por ello que no retiene el violeta cristal y por esto las células se tiñen con safranina y las observamos roja
Formas y agrupaciones bacteriana
Los principales tipos de formas bacterianas son:
*Cocos (células más o menos esféricas)
*Bacilos (en forma de bastón, alargados),que a su vez pueden tener varios aspectos:cilíndricos,fusiformes y en forma de maza, etc.
Atendiendo a los tipos de extremos, éstos pueden ser:
*Redondeados (lo más frecuente)
*Cuadrados
*Biselados
*Afilados
*Espirilos: al igual que los bacilos, tienen un eje más largo que otro, pero dicho eje no es recto, sino que sigue una forma de espiral, con una o más de una vuelta de hélice.
*Vibrios: proyectada su imagen sobre el plano tienen forma de coma, pero en el espacio suelen corresponder a una forma espiral con menos de una vuelta de hélice.
Otros tipos de formas son filamentos, ramificados o no,anillos casi cerrados, y formas con prolongaciones (con prostecas).
2011-0481
ResponderEliminarFAMILIAS DE VIRUS.
-PicoRNAvirus:
Por picornavirus se conoce a todo virus que pertenece a la familia Picornaviridae. Los picornavirus son virus no envueltos, de única cadena ARN con cápside icosaédrica.
Su genoma es inusual ya que tiene una proteína en el extremo 5 ‘que se utiliza como cebador para la transcripción por la ARN polimerasa. El nombre se deriva de “pico”, que significa pequeño, ya que es literalmente un pequeño virus de ARN.
Los picornavirus se separan en una serie de géneros e incluyen muchos importantes patógenos de humanos.
Las enfermedades que causan son muy variadas, que van desde aguda “gripe común” a enfermedades como la poliomielitis, y otras infecciones crónicas.
- Los Calicivirus:
Son Virus RNA pequeños pertenecientes a la familia Caliciviridae. Dentro de esta familia destacan dos géneros: los Norovirus, también conocidos como "virus tipo Norwalk" o "virus pequeños redondos estructurados" y los Sapporovirus.
- Hepadnavirus:
Los hepadnavirus son un subgrupo de los virus que producen hepatitis.
Es uno de los virus más importantes desde el punto de vista médico. Infecta a más de un cuarto de billón de personas y es una de las principales causas de cirrosis y cáncer hepático (carcinoma hepatocelular). HBV, también representa el primer ejemplo del éxito de una vacuna recombinante para una enfermedad contagiosa humana, y puede tener importantes implicaciones para otras enfermedades de transmisión sexual, como el HIV.
- Reovirus:
Grupo de tres virus de doble cadena de ARN presentes en todas partes, identificados en los aparatos respiratorio y digestivo tanto de personas sanas como de enfermas. Los reovirus han sido implicados en algunos casos de enfermedades de vías respiratorias superiores y de gastroenteritis infantiles.
- Los Rhabdovirus:
Son virus con cápside helicoidal. Incluye al virus de la rabia y de la estomatitis vesicular (que es el organismo modelo). El virus de la estomatitis vesicular es un virus de 180x75nm, con cápsida rodeada por una envoltura con proyecciones constituídas por la proteína G.
- Togavirus:
Familia de arbovirus a la que pertenecen los organismos causantes de encefalitis, del dengue, de la fiebre amarilla y de la rubéola.
- Bunyavirus:
Género de virus ARN de vertebrados de la familia Bunyaviridae.
-Papovavirus:
Grupo de pequeños virus ADN, algunos de los cuales pueden ser potencialmente productores de cáncer. La verruga humana está producida por un tipo de papovavirus. Algunos tipos de papovavirus son: papovavirus del papiloma, papovavirus del polioma y papovavirus SV.
- Paramixovirus:
Miembro de una familia de virus que abarca a los organismos causantes de la gripe, de las paperas y de algunas infecciones respiratorias.
- Retrovirus:
Virus de ácido ribonucleico que posee una enzima que le permite convertir su material genético en ácido desoxirribonucleico cuando ha penetrado en una célula viva. Al grupo de los retrovirus pertenece el virus del sida.
- Filovirus:
Género de virus con ácido ribonucleico de cadena simple con sentido negativo, de la familia Filoviridae. El género incluye los virus Ébola y Marburg.
- Poxvirus:
Miembro de una familia de virus que incluye a los organismos que producen el molusco contagioso, la viruela y la vacuna.
*DIAGNOSTICO DE LAS VIROSIS.
¿Cómo infectan los virus al ser humano?
Los virus pueden penetrar en el cuerpo humano por cualquiera de sus “puertas de entrada,” pero por lo general entran por la nariz y la boca. Una vez dentro del cuerpo, el virus se adhiere a la superficie de la célula que piensa atacar, denominada célula huésped.
Una vez dentro, comienza a elaborar otros virus idénticos a partir de la proteína de la célula huésped. Los nuevos virus pueden salir a través de la membrana externa de la célula huésped, a la que a veces destruyen antes de abandonarla, y se lanzan al ataque contra otras células huésped. El proceso prosigue hasta que el cuerpo elabora suficiente cantidad de anticuerpos y otras defensas para derrotar a los virus invasores.
yuleika veloz 2011-0481
ResponderEliminarBIONES ACELULARES
El término acelular significa literalmente "sin células". Se aplica principalmente a los virus, pero también a provirus y viroides, que son entidades biológicas no formadas por células, a diferencia de bacterias, compuestas por células procariotas, y de hongos, vegetales y animales, formados por células eucariotas.
LOS VIROIDES I y II
son agentes infecciosos que, al igual que los virus, tienen un ciclo extracelular que se caracteriza por la inactividad metabólica y unciclo intracelular en el que causan infección al huésped susceptible, pero que a diferencia de los virus, los viroides no poseen proteínas ni lípidos y están constituidos por una cadena cíclica corta de ARN, circular o con forma de varilla, (que no codifica proteínas).1 Es importante decir que tanto su forma intracelular como extracelular son las mismas (ARN desnudo), los mecanismos por los cuales éstos logran causar infección están relacionados con la autocatálisis de su material genético.
Estas pequeñas moléculas de ARN circular de cadena sencilla formadas por unos pocos cientos de nucleótidos. Carecen de cápsida y su tamaño es una milésima parte del de los virus más pequeños.
- Aparecen con mayor abundancia en el núcleo de las células infectadas.
- Sólo han sido detectados en plantas, en las que producen una gran variedad de enfermedades y pueden ser transmitidos tanto horizontal como verticalmente.
- No existen ninguna evidencia de que los viroides son traducidos a proteínas, ni se conoce cómo causan las enfermedades. Lo que sí se sabe es que son replicados por los enzimas del huésped.
- Algunas semejanzas entre la secuencia de nucleótidos de los viroides y ciertos secuencias del genoma de las plantas
no traducidas (intrones) sugieren que los viroides evolucionaron a partir de los intrones.
YULEIKA VELOZ 2011-0481
ResponderEliminarFARMACOS ANTIVIRALES
GENERALIDADES
Actualmente se dispone de farmacos antivirales de probada eficacia terapeutica y profilactica para varias infecciones virales comunes y algunas potencialmente mortales. Ademas, continua la busqueda de nuevos farmacos antivirales y nuevas estrategias terapeuticas para el tratamiento de las enfermedades virales.
Se pueden clasificar en tres grandes grupos a los agentes quimioterapicos activos frente a los virus:
Farmacos Viricidas: farmacos que directamente inactivan los virus intactos.
Detergentes.
Disolventes organicos: eter, cloroformo.
Luz ultravioleta.
Podofilinas, las cuales destruyen ademas tejidos del huesped.
Farmacos Antivirales: farmacos que inhiben procesos especificos del virus, como la union a la celula, la decapsidacion del genoma viral o el ensamblaje de los nuevos virus, o, preferentemente, inhiben la sintesis macromolecular dirigida por el virus.
Farmacos Inmunomodulares: comprenden los que reemplazan las respuestas inmunitarias deficientes del huesped y los que favorecen a los endogenos.
ANTIVIRALES QUE IMPIDEN LA ENTRADA DEL VIRUS A LA CELULA
Amantadina: actua sobre el virus influenza A. Su efecto principal es inhibir la decapsidacion viral o el desensamblaje del virion durante la endocitosis.
Rimantadina: derivado 4-10 veces mas activa que la amantadina.
Oseltamivir: inhibe la actividad de neuraminidasa del virus influenza.
Zanamivir:inhibe la actividad de neuraminidasa del virus influenza.
Docosanol: interfiere en la fusion del VHS-1 y VHS-2 a las membranas de las celulas del huesped. Es de uso topico.
ANTIVIRALES QUE ACTUAN SOBRE LA FASE DE REPLICACIÓN DEL GENOMA VIRAL
Aciclovir: inhibe la sintesis del ADN viral. Tiene accion sobre los herpesvirus.
Valaciclovir: profarmaco del Aciclovir.
Ganciclovir: inhibe la sintesis del ADN viral. Tiene accion sobre los herpesvirus, pero su caracteristica singular es una potente inhibicion de la replicacion del CMV.
Valganciclovir: profarmaco del Ganciclovir.
Adefovir: Actua como inhibidor competitivo de las polimerasas del ADN y transcriptasas inversas virales con respecto al dATP. Se selectividad se relaciona con una mayor afinidad por la ADN polimerasa del VHB que por las polimerasas de las celulas huesped. Esta aprobado para el tratamiento de la hepatitis B cronica.
Cidofovir: inhibe la sintesis de ADN viral de los herpesvirus.
Penciclovir: inhibe la sintesis de ADN viral de los herpesvirus.
Famciclovir. Profarmaco del Penciclovir de administracion oral.
Fomivirseno. Inhibe la replicacion del CMV humano. Se administra de forma intravitrea.
Foscarnet: actua sobre los herpesvirus y la transcriptasa reversa del HIV. Es inhibidor frente a la mayoria de las cepas de CMV resistentes a ganciclovir y de VHS y VVZ resistentes a aciclovir.
Idoxuridina. Inhibe la sintesis de ADN viral. Actua en especial sobre los herpesvirus y poxvirus.
Ribavirina: se utiliza en el tratamiento de la infección por HCV y por el VSR.
Trifluridina.
Vidarabina
RETROVIRALES
Retroviridae es una familia de virus que comprende los retrovirus. Son virus con envoltura que presentan un genoma de ARN monocatenario de polaridad + y se replican de manera inusual a través de una forma intermedia de ADN bicatenario. Este proceso se lleva a cabo mediante una enzima: la retrotranscriptasa o transcriptasa inversa, que dirige la síntesis de ADN a través de ARN y posee una importancia extraordinaria en la manipulación genética. Una vez se ha pasado de ARN monocatenario a ADN se inserta dentro del ADN propio de la célula infectada donde se comporta como un gen más (véase Ciclo reproductivo de los virus).1 Por tanto, se incluyen en el Grupo VI de la Clasificación de Baltimore.
2011-0175 Jafre Puello
ResponderEliminardiapositiva # 1
virologia
Virología es el estudio de virus y virus como agentes: clasificación y evolución, su estructura, sus formas de infectar y explotar las células para la reproducción del virus, las enfermedades que causan, las técnicas para aislar y la cultura de ellos y su uso en la investigación y terapia.
Virología es considerada una parte de la microbiología o de patología.
Una importante rama de la Virología es la clasificación de virus. Virus pueden clasificarse según la célula huésped que infectan: virus animales, virus de plantas, hongos virus y bacteriófagos (virus que infectan bacterias, que son los virus más complejos).
Otra clasificación utiliza la forma geométrica de su cápside (a menudo una hélice o un icosaedro) o la estructura del virus (por ejemplo, la presencia o la ausencia de un envolvente de lípidos). Gama de virus en tamaño de unos 30 nm a aproximadamente 450 nm, lo que significa que la mayoría de ellos no puede verse con microscopios de luces.
La forma y la estructura del virus ha sido estudiado por cristalografía de rayos X, microscopía electrónica y RMN.
El sistema de clasificación más útil y más ampliamente utilizado distingue virus segun el tipo de ácido nucleico que utilizan como material genético y el método de replicación viral que emplean coaxial células huésped en la producción de más virus:
Virus de ADN (divididas en virus de ADN bicatenario y los virus de ADN monocatenario mucho menos común),
Virus ARN (dividieron en virus de ARN monocatenario de sentido positivo, virus de ARN monocatenario de sentido negativo y los virus de ARN bicatenario mucho menos común),
invertir transcribir virus (virus de ADN de transcripción inversa de bicatenario y monocatenario transcripción inversa RNA virus incluyendo retrovirus).
Mat:2011-0289 -VIROLOGIA-
ResponderEliminarLa virología (rama de la microbiologia) es el estudio de los virus: su estructura, clasificación y evolución, su manera de infectar y aprovecharse de las células huésped para la reproducción del virus, su interacción con los organismos huéspedes, su inmunidad, la enfermedad que causan, las técnicas para su aislamiento, cultivo y su uso en investigación y terapia. La virología es considerada un sub-campo de la microbiología y la medicina. La virologia también es una rama taxonómica de la biología para estudiar los virus y todos sus componentes.
Su investigación incluye:
la replicación viral
los patógenos virales.
la inmunología viral.
las vacunas virales.
los métodos de diagnóstico.
la quimioterapia antiviral.
las medidas de control de una infección.
los diferentes signos que manifiestan la presencia de virus.
Mat:2011-0289 -ESTRUCTURA DE LOS VIRUS-
ResponderEliminarLos virus presentan una amplia diversidad de formas y tamaños, llamadas «morfologías». Son unas 100 veces más pequeños que las bacterias. La mayoría de los virus estudiados tienen un diámetro de entre 10 y 300 nanómetros. Algunos Filovirus tienen un tamaño total de hasta 1.400 nm, sin embargo, sólo miden unos 80 nm de diámetro.71 La mayoría de virus no pueden ser observados con un microscopio óptico, de manera que se utilizan microscopios electrónicos de barrido y de transmisión para visualizar partículas víricas.72 Para aumentar el contraste entre los virus y el trasfondo se utilizan tinciones densas en electrones. Son soluciones de sales de metales pesados como wolframio, que dispersan electrones en las regiones cubiertas por la tinción. Cuando las partículas víricas están cubiertas por la tinción (tinción positiva), oscurecen los detalles finos. La tinción negativa evita este problema, tiñendo únicamente el trasfondo.73
Una partícula vírica completa, conocida como virión, consiste en un ácido nucleico rodeado por una capa de protección proteica llamada cápside. Las cápsides están compuestas de subunidades proteicas idénticas llamadas capsómeros.74 Los virus tienen un «envoltorio lipídico» derivado de la membrana celular del huésped. La cápside está formada por proteínas codificadas por el genoma vírico, y su forma es la base de la distinción morfológica.75 76 Las subunidades proteicas codificadas por los virus se autoensamblan para formar una cápside, generalmente necesitando la presencia del genoma viral. Sin embargo, los virus complejos codifican proteínas que contribuyen a la construcción de su cápside. Las proteínas asociadas con los ácidos nucleicos son conocidas como nucleoproteínas, y la asociación de proteínas de la cápside vírica con ácidos nucleicos víricos recibe el nombre de nucleocápside.77 78 En general, hay cuatro tipos principales de morfología vírica:
Helicoidal
las cápsides helicoidales se componen de un único tipo de capsómero apilado alrededor de un eje central para formar una estructura helicoidal que puede tener una cavidad central o un tubo hueco. Esta formación produce viriones en forma de barra o de hilo, pueden ser cortos y muy rígidos, o largos y muy flexibles. El material genético, normalmente ARN monocatenario, pero a veces ADN monocatenario, queda unido a la hélice proteica por interacciones entre el ácido nucleico con carga negativa y la carga positiva de las proteínas. En general, la longitud de una cápside helicoidal está en relación con la longitud del ácido nucleico que contiene, y el diámetro depende del tamaño y la distribución de los capsómeros. El conocido virus del mosaico del tabaco es un ejemplo de virus helicoidal.79
Icosaédrica
la mayoría de virus que infectan los animales son icosaédricos o casi-esféricos con simetría icosaédrica. Un icosaedro regular es la mejor manera de formar una carcasa cerrada a partir de subunidades idénticas. El número mínimo requerido de capsómeros idénticos es doce, cada uno compuesto de cinco subunidades idénticas. Muchos virus, como los rotavirus, tienen más de doce capsómeros y parecen esféricos, manteniendo esta simetría. Los ápices de los capsómeros están rodeados por otros cinco capsómeros y reciben el nombre de pentones. Las caras triangulares de éstos también se componen de otros seis capsómeros y reciben el nombre de hexones.
Envoltura
algunas especies de virus se envuelven en una forma modificada de una de las membranas celulares, o bien es la membrana externa que rodea una célula huésped infectada, o bien membranas internas como la membrana nuclear o el retículo endoplasmático, consiguiendo así una bicapa lipídica exterior conocida como envoltorio vírico. Esta membrana se rellena de proteínas codificadas por el genoma vírico y el del huésped, la membrana lipídica en sí y todos los carbohidratos presentes son codificados completamente por el huésped. El virus de la gripe y el VIH utilizan esta estrategia. La mayoría de virus envueltos dependen de la envoltura para infectar.81
Mat:2011-0289 FAMILIAS DE VIRUS
ResponderEliminarI- Adenoviridae
Mastadenovirus
Es un taxón que afecta a vertebrados.
Aden-(var.”adeno-“) Elemento prefijo del gr. “adén”, que significa “glándula”.
Viriones isométricos no envueltos.
Nucleocapside angular 70-90 nm diámetro.
Simetría icosaedrica.
II- Virus de la peste porcina africana
Infecta a cerdos domesicos, facoceros, potamóquero de río y garrapatas.
Causa fiebres hemorrágicas.
La peste africana se limitaba al continente que lleva su nombre hasta 1957 cuando la enfermedad se informó en otros países.
III- Arenaviridae
Virus que afecta a vertebrados.
Arena, viene del latín con la familia de términos del árido.
Virus envueltos, ligeramente pleomórficos; esféricas. Viriones probablemente contengan 2 nucleocapsides envueltas.
Causantes de fiebres hemorrágicas.
IV- Artevivirus
Familia de virus que infectan animales.
Arterivirus: arteritis. Inflamación vasos sanguíneos.
Estos virus son pequños, con envoltura y con forma icosaédrica.
V- Astroviridae
Virus que afecta a vertebrados.
Astro- es un prefijo lo cual significa estrellas.
Human astrovirus: Virones no envueltos. Nucleocapside isométrica. 27-30 nm diámetro. Simetría polihedrica. Con vértices.
VI- Baculoviridae
Virus que infecta vertebrados. Báculo: del latín, “palo”. Refiriéndose a la forma nucleocápside.
Son visibles en secciones de tejido infectado. Viriones ocluidos por cristales de proteínas o no ocluidos. Virion envuelto y pleomorfo forma de vara. Nucleocapsides en forma de vara. 200-450 nm long. Y 30-100 de diámetro.
VII- Badnavirus
Afecta a plantas
Etimología: Baciliforme Ba, DNA.
VIII- Barnaviridae
Infecta a hongos.
Etimología: Baciliforme Ba, RNA.
Sin envoltura, con un baciliformes T=1 simetría icosaédrica, aproximadamente 50 nm de longitud y 20 nm de gran tamaño.
IX- Birnaviridae
Afecta a vertebrados.
Birna: para el ARN bipartito.
No envueltos, de 60 nm de diámetro, virión icosaédricO con T=13 simetría.
X- Bromoviridae
Afecta a plantas.
Bromo- porque infecta a Bromus inermis.
Viriones no envueltos, icosaédrico o baciliformes 26-35 nm de diámetro.
XI- Bunyaviridae
Afecta a vertebrados y a plantas.
Viriones pleomorficos, esferoidales.
“Bunyamwera” ciudad de Uganda donde se aisló.
Envueltos, esféricos. Diámetro de 80 a 120 nm.
XII- Caliciviridae
Virus que infecta a vertebrados y a invertebrados.
“Calici” porque presenta depresiones calciformes.
Viriones no están envueltos y la capside es isométrica. Simetría icosaedrica.
XIII- Capillovirus
Infecta a plantas.
Del latín “capella”, diminutivo de capa.
El 3 es poliadenilado. Codifica por lo menos 3 proteínas.
Mat:2011-0289
ResponderEliminarDiagnostico de la Virosis
Los virus son organismos tan pequeños, que su tamaño les permite pasar a través de los filtros que retienen bacterias. Las características básicas para identificar a un virus son: la composición del ácido nucleico, la disposición de la cápside, la presencia o ausencia de una envoltura, el tamaño del virus y el huésped.
MUESTRAS PARA DIAGNOSTICO:
Las muestras deben tomarse precozmente en la fase aguda de la infección. Deben transladarse al laboratorio a la mayor brevedad posible, en medio de transporte. Las muestras más comunes son:
• Torundas nasofaríngeas: adecuados para el diagnóstico de enterovirus, adenovirus, y herpes simplex. Las nasofaringeas se prefieren para el virus respiratorio sincicial y la mayoria de los otros respiratorios. Las nasales son las mejores para los rinovirus.
• Aspirados faríngeos: son mejores que los hisopos (torundas) nasofaringeos, pero estos últimos son mas prácticos.
• Torundas rectales y muestras de heces: Aquí se muestran virus no cultivables como rotavirus, o adenovirus, que se detectan por microscopía electrónica o con técnicas de detección de antígenos.
• Secreciones, Sangre o Líquido cefalorraquídeo.
• En algunas virosis excepcionalmente el diagnóstico solo puede efectuarse en Biopsia de cerebro, como en la encefalitis por herpes simplex; en Huéspedes inmunodeprimidos y con citomegalovirus, el virus solo puede identificarse en el órgano afectado.
Mat:2011-0289 -ETAPAS DE LA REPLICACION VIRAL-
ResponderEliminarAl carecer de las enzimas biosintéticas necesarias para su replicación, los virus no pueden replicarse en medios de cultivo inertes sino que necesitan de células vivas. La replicación se realiza en el interior de una célula huésped susceptible.
En los comienzos de la virología, se utilizaron embriones de pollo o pato y/o animales de experimentación para replicar los virus. Posteriormente, con el advenimiento de las técnicas para mantener vivas células “in vitro” se pudieron desarrollar numerosos cultivos celulares, aptos para la replicación de la mayoría de los virus que causan patología humana y animal. En la actualidad, para el aislamiento de virus se utilizan casi exclusivamente los cultivos celulares.
Luego de la penetración del virus en una célula susceptible, la replicación se lleva a cabo utilizando los mecanismos biosintéticos de la célula, dirigidos por la información contenida en el ácido nucleico viral. Este actúa como codificador de los mecanismos enzimáticos de síntesis de ácidos nucleicos y proteínas. De esta forma, la célula parasitada por un virus e vez de producir sus propios ácidos nucleicos y proteínas, recibe órdenes codificadas en el genoma viral parta sintetizar productos virales que, luego de su ensamble, darán lugar a la formación de nuevos viriones. Estos saldrán de la célula donde replicaron para iniciar nuevos ciclos de replicación en otras células susceptibles.
La replicación viral puede dividirse en las siguientes fases:
Adsorción: que depende de la interacción de proteínas de superficie del virus con receptores celulares específicos. Por ejemplo el receptor CD4 del linfocito T y la glicoproteína gp120 del virus HIV, las hemaglutininas de los virus influenza y los receptores de ácido acetil-neuramínico en la superficie de las células epiteliales, etc.
Penetración y decapsidación: la penetración a través de la membrana plasmática de la célula puede efectuarse por los siguientes mecanismos: por pasaje directo, por fusión de la membrana celular con la envoltura viral, o por endocitosis o viropexis, mecanismo más frecuente, es un fenómeno similar a la fagocitosis, en el cual el virus es incluído en una vacuola fagocítica. Luego de la penetración se produce la decapsidación, es decir la liberación del ácido nucleico viral de la nucleocápside.
Eclipse: en esta etapa no pueden observarse viriones en el interior de la célula, ni se recupera virus infeccioso.
Latencia: este período va desde que se pierde la infectividad inicial hasta que se la recupera debido a la aparición de los nuevos viriones. Durante las etapas de eclipse y latencia ocurre la biosíntesis de los ácidos nucleicos y de las proteínas virales.
Maduración: de produce el ensamble de los ácidos nucleicos neoformados con las proteínas de la cápside.
Liberación: los nuevos viriones salen de la célula por lisis de la misma, como el virus polio; o bien por brotación a través de la membrana celular, como los paramixovirus o arenavirus; o de la membrana nuclear, como los herpesvirus o del retículo endoplásmico, como los togavirus
Mat:2011-0289 -ADHESION-
ResponderEliminarLa adhesión es la propiedad de la materia por la cual se unen y plasman dos superficies de sustancias iguales o diferentes cuando entran en contacto, y se mantienen juntas por fuerzas intermoleculares.
La adhesión ha jugado un papel muy importante en muchos aspectos de las técnicas de construcción tradicionales. La adhesión del ladrillo con el mortero (cemento) es un ejemplo claro.
Mecanismos de adhesión:
La cohesión es la causa de que el agua forme gotas, la tensión superficial hace que se mantengan esférica y la adhesión la mantiene en su sitio.
Las gotas de agua son más planas sobre la flor de Hibiscus ya que tienen mejor adhesión.
Cinco mecanismos han sido propuestos para explicar por qué un material se adhiere a otro.
Adhesión mecánica
Los materiales adhesivos rellenan los huecos o poros de las superficies manteniendo las superficies unidas por enclavamiento. Existen formas a gran escala de costura, otras veces a media escala como el velcro y algunos adhesivos textiles que funcionan a escalas pequeñas. Es un método similar a la tensión superficial
Adhesión química
Dos materiales pueden formar un compuesto químico al unirse. Las uniones más fuertes se producen entre átomos donde hay permutación (enlace iónico) o se comparten electrones (enlace covalente). Un enlace más débil se produce cuando un átomo de hidrógeno que ya forma parte de una partícula se ve atraída por otra de nitrógeno, oxígeno o flúor, en ese caso hablaríamos de un puente de hidrógeno.
Adhesión dispersiva
En la adhesión dispersiva, dos materiales se mantienen unidos por las fuerzas de van der Waals: la atracción entre dos moléculas, cada una de las cuales tiene regiones de carga positiva y negativa. En este caso, cada molécula tiene una región de mayor carga positiva o negativa que se une a la siguiente de carga contraria. Este efecto puede ser una propiedad permanente o temporal debido al movimiento continuo de los electrones en una región.
En la ciencia de superficies el término "adhesión" siempre se refiere a una adhesión dispersiva. En un sistema sólido-líquido-gas normal (como una gota de un líquido sobre una superficie rodeada de aire) el ángulo de contacto es usado para cuantificar la adhesividad. En los casos donde el ángulo de contacto es bajo la adhesión está muy presente. Esto se debe a que una mayor superficie entre el líquido y el sólido conlleva una energía superficial mayor.
Adhesión electrostática
Algunos materiales conductores dejan pasar electrones formando una diferencia de potencial al unirse. Esto da como resultado una estructura similar a un condensador y crea una fuerza electrostática atractiva entre materiales. mi con diversos materiales les pueden ayudar
Mat:2011-0289
ResponderEliminarPenetración y decapsidación: la penetración a través de la membrana plasmática de la célula puede efectuarse por los siguientes mecanismos: por pasaje directo, por fusión de la membrana celular con la envoltura viral, o por endocitosis o viropexis, mecanismo más frecuente, es un fenómeno similar a la fagocitosis, en el cual el virus es incluído en una vacuola fagocítica. Luego de la penetración se produce la decapsidación, es decir la liberación del ácido nucleico viral de la nucleocápside.
Penetración de los Bacteriófagos:
Inyección del material genético viral: Después de la adsorción, se produce un cambio configuracional en las proteínas de la placa basal, alguna de las cuales tienen actividad enzimática y producen un poro en la membrana citoplasmática de la célula. La vaina del fago se contrae y el material genético viral ingresa en la célula, mientras que la envoltura proteica queda en el exterior.
Para entrar en una célula, los fagos se acoplan a receptores específicos en la superficie de la bacteria, que pueden ser lipopolisacáridos, ácidos teicoicos, proteínas o incluso flagelos. Por ello, cada fago solo podrá infectar ciertas bacterias según sus receptores. Puesto que los fagos no son móviles, dependen de encuentros al azar con los receptores adecuados en solución para poder infectar una bacteria.
Parece que los bacteriófagos presentan una especie de jeringa mediante la cual introducen su material genético en el interior de la célula. Tras el reconocimiento del receptor adecuado, la cola y cuello del fago se contraen, quedando así el fago acoplado a la superficie celular. El material genético puede ser ahora introducido a través de la membrana o bien simplemente depositado sobre la superficie. No se descarta que pueda haber fagos con otros métodos diferentes para introducir su material genético en la célula.
partes 3 y 4 dios te bendiga 2011-0119
ResponderEliminarLA CÉLULA BACTERIANA
Las bacterias son células muy sencillas; carecen de núcleo y tampoco presentan orgánulos en el citoplasma. Se las denomina Procariotas. Son organismos unicelulares y se encuentran en todos los ecosistemas.
Las bacterias son un numeroso grupo de seres vivos, con características muy diversas. En la clasificación de los Dominios, Woese, aparecen dos grupos de Procariotas:
el Dominio Archaea, que engloba a los organismos más antiguos del Planeta, y
el Dominio Bacteria, en el que se encuentran la gran mayoría de los organismos bacterianos actuales, también conocidos con el nombre de Eubacterias.
Las bacterias se caracterizan por no tener núcleo patente, ni clorofila típica, aunque pueden poseer otros pigmentos similares como bacterioclorina, bacterioclorofila y bacteriopurpurina, y sin pseudópodos. La bacterioclorina es un pigmento verde fotosintetizante que, según ciertos autores, acompaña a la bacteriopurpurina y a la bacterioclorifila en las algas purpúreas.
Tamaño de los Virus
Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus.
Su pequeño tamaño explica lo tardío del descubrimiento de estos agentes. La primera referencia sobre la existencia de los virus se debe al botánico ruso Dimitri Ivanovski en 1892. Este investigador buscaba el agente causante de la enfermedad denominada mosaico del tabaco, y llegó a la conclusión de que debía tratarse de una toxina o de un organismo más pequeño que las bacterias, pues el agente atravesaba los filtros que retenían las bacterias. Denominó a estos agentes patógenos virus filtrables.
Cristian Rafael Guerrero Mendoza Mat 89275
ResponderEliminarIntroducción
Virología
La virología (rama de la microbiologia) es el estudio de los virus: su estructura, clasificación y evolución, su manera de infectar y aprovecharse de las células huésped para la reproducción del virus, su interacción con los organismos huéspedes, su inmunidad, la enfermedad que causan, las técnicas para su aislamiento, cultivo y su uso en investigación y terapia. La virología es considerada un sub-campo de la microbiología y la medicina. La virologia también es una rama taxonómica de la biología para estudiar los virus y todos sus componentes.
Su investigación incluye:
la replicación viral
los patógenos virales.
la inmunología viral.
las vacunas virales.
los métodos de diagnóstico.
la quimioterapia antiviral.
las medidas de control de una infección.
los diferentes signos que manifiestan la presencia de virus.
Cristian Rafael Guerrero Mendoza Mat 89275
ResponderEliminarEstructura General de los Virus
Un virus está compuesto de una molécula de ácido nucleico y una envoltura proteínica. Ésta es la estructura básica de un virus, aunque algunos de ellos pueden añadir a esto la presencia de alguna enzima, bien junto al ácido nucleico, como la transcriptasa inversa de los retrovirus, bien en la envoltura, para facilitar la apertura de una brecha en la membrana de la célula hospedadora.
A la unidad formada por el ácido nucleico y la envoltura proteínica se le denomina tambiénvirión.
El ácido nucleico es solamente de un tipo, ADN o ARN, nunca los dos.
Atendiendo al tipo de ácido nucleico se distinguen cuatro clases de virus:
- ADN de cadena doble
- ADN de cadena sencilla
- ARN de cadena doble
- ARN de cadena sencilla
La envoltura proteínica recibe el nombre de cápsida. Está formada por unas subunidades idénticas denominadas capsómeros. Los capsómeros son proteínas globulares que en ocasiones tienen una parte glicídica unida. Se ensamblan entre sí dando a la cubierta una forma geométrica.
Atendiendo la forma de la cápsida, se pueden distinguir los siguientes tipos de virus:
- Cilíndricos o helicoidales: los capsómeros, que son de un solo tipo, se ajustan entorno una hélice simple de ácido nucleico. Un ejemplo lo constituye el virus del mosaico del tabaco.
- Icosaédricos: los capsómeros, que suelen ser de varios tipos, se ajustan formando un icosaedro regular (es decir, 20 caras triangulares y 12 vértices), y dejando un hueco central donde se sitúa el ácido nucleico fuertemente apelotonado. Algunos forman poliedros con más caras que el icosaedro, y algunos presentan fibras proteicas que sobresalen de la cápsida.
Un ejemplo lo constituyen los adenovirus, entre los que se encuentran los virus de los resfriados y faringitis.
Cristian Rafael Guerrero Mendoza Mat 89275
ResponderEliminarVIROLOGIA. SIMETRIA VIRAL
La simetría es la forma que adopta un virus en el espacio y está dad por la estructura de su nucleocápside. La simetría puede ser helicoidal, icosaédrica o compleja.
Los virus con simetría helicoidal presentan una nucleocápside cilindrica que puede estar extendida como en el virus del mosaico del tabaco. Este es un virus desnudo. En otros casos la nucleocápside está arrollada sobre sí misma y recubierta por una envoltura, como en el
virusInfluenza.
Los virus con simetría icosaédrica son poliedros regulares con 20 caras triangulares, 30 aristas y 12 vértices. Son ejemplos de virus con simetría icosaédrica desnudos el virus de la poliomielitis o losadenovirus. Los herpesvirus y togavirus son virus con simetría icosaédrica y recubiertos por una envoltura.
Se denominan virus de simetría compleja a aquellos que presentan una nucleocápside helicoidal o icosaédrica recubierta por una envoltura laxa. Ejemplos de éstos virus son los poxvirus (forma de ladrillo), los rhabdovirus (forma de bala) y algunos bacteriófagos que poseen una cabeza con simetría icosaédrica y una cola con simetría helicoidal.
Simetría cúbica: Toda la simetría cúbica observada en los virus de los animales, hasta la fecha, es del patrón icosaédrico, la disposición más eficiente de subunidades en una cubierta cerrada. Conocer las reglas que orientan la simetría icosaédrica hace posible determinar el número de capsómeros en una partícula, característica importante de la clasificación de los virus.
El ácido nucleico viral se condensa dentro de las partículas isoméricas; en la condensación del ácido nucleico en una forma adecuada para su acumulación participan proteínas "nucleares" codificadas por el virus o, en el caso de los papovirus, histonas celulares. No han podido definirse aún las reglas para la incorporación del ácido nucleico en las partículas isométricas; probablemente una "secuencia de empaque" participa en el ensamble, aunque en general no son cruciales las estructuras primarias, secundarias y terciarias del ácido nucleico. Existen limitaciones de tamaño en las moléculas de ácido nucleico que se pueden empacar o acumular en una cápside icosaédrica determinada. Las cápsides icosaédricas se forman de manera independiente del ácido nucleico. La mayor parte de los preparados de virus isométricos contendrán algunas partículas "vacías", desprovistas de ácido nucleico viral. Ejemplos de simetría cúbica son los grupos de virus tanto de ADN como de ARN.
Simetría helicoidal: En los casos de simetría helicoidal, las subunidades proteínicas están fijas de manera periódica al ácido nucleico viral, con lo que lo amplían en una espiral. El complejo filamentoso de proteína y ácido nucleico viral (nucleocápside) se enrolla a continuación dentro de una cubierta que contiene lípidos. Por lo tanto, a diferencia del caso de las estructuras icosaédricas, existe una interacción periódica regular entre la proteína de la cápside y el ácido nucleico en el virus que tiene simetría helicoidal. No es posible que se formen partículas helicoidales "vacías".
Cristian Rafael Guerrero Mendoza Mat 89275
ResponderEliminarTamaño de los Virus
Tamaño
Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus.
Su pequeño tamaño explica lo tardío del descubrimiento de estos agentes. La primera referencia sobre la existencia de los virus se debe al botánico ruso Dimitri Ivanovski en 1892. Este investigador buscaba el agente causante de la enfermedad denominada mosaico del tabaco, y llegó a la conclusión de que debía tratarse de una toxina o de un organismo más pequeño que las bacterias, pues el agente atravesaba los filtros que retenían las bacterias. Denominó a estos agentes patógenos virus filtrables.
En 1897, el microbiólogo holandés Martinus Beijerink realizó experimentos similares a los de Ivanovski, y llegó a desechar la idea de las toxinas, pues se trataba de un agente capaz de reproducirse, ya que mantenía su poder infeccioso de unas plantas a otras, sin diluirse su poder patógeno. Poco después, los microbiólogos alemanes Frederick Loeffler y Paul Frosch descubrieron que la fiebre aftosa del ganado era producida por un virus filtrable que actuaba como un agente infeccioso.
En la década de los 30, con el uso de filtros de tamaño de poro inferior, con las técnicas de cultivo celular in vitro que permitían la obtención de gran cantidad de virus, con la ultracentrifugación y finalmente con el microscopio electrónico y la difracción de rayos X, se logró visualizar a estos agentes.
Los virus son cristalizables, como demostró W. Stanley en 1935. Esto depende del hecho de que las partículas víricas tienen formas geométricas precisas y que son idénticas entre sí, lo cual las separa de la irregularidad característica de los organismos, las células o los orgánulos, y las acerca a las características de los minerales y de agregados de macromoléculas como los ribosomas. Al tener un volumen y forma idénticos, las partículas víricas tienden a ordenarse en una pauta tridimensional regular, periódica, es decir, tienden a cristalizar.
Cristian Rafael Guerrero Mendoza Mat 89275
ResponderEliminarFamilia de los Virus
1- Picornaviridae: es una familia de virus infectivos para animales. Contienen un genoma ARN monocatenario positivo y por lo tanto se incluyen en el Grupo IV de la Clasificación de Baltimore. El genoma ARN es inusual porque tiene una proteína en el terminal 5' que se utiliza como iniciador de la transcripción por la ARN polimerasa. El nombre se deriva de "pico", que significa pequeño, con lo que "picornavirus" significa literalmente "virus RNA pequeños".
2- Caliciviridae: es una familia de virus infectivos para animales y causantes de gastroenteritis en humanos. Los calicivirus han sido encontrados en la mayoría de los animales domésticos y muchos silvestres, como cerdos, conejos, gallinas y anfibios. Contienen un genoma ARN monocatenario positivo y por lo tanto se incluyen en el Grupo IV de la Clasificación de Baltimore.
3- Hepadnaviridae es una familia de virus que causan infecciones en hígado de humanos y de animales. Los hepadnavirus tienen un genoma muy corto de ADN parcialmente de doble hélice y parcialmente de hélice simple, circular. Como su replicación involucra un ARN intermedio, se incluyen en el Grupo VII de la Clasificación de Baltimore.
4- Reoviridae es una familia de virus que pueden afectar el sistema gastrointestinal (tales como rotavirus ) y tracto respiratorio . Los virus de la familia Reoviridae han genomas que consiste en segmentar, ARN de doble cadena (dsRNA). El nombre "Reoviridae" se deriva de las vías respiratorias entéricas virus huérfanos.
5- Rhabdoviridae es una familia de virus infectivos para animales y plantas. Entre los animales infectados están insectos, peces y mamíferos, incluidos los humanos. Estos virus presentan una forma alargada parecida a una bala. Poseen un genoma de ARN monocatenario de sentido negativo por lo que se incluye en el Grupo V de la Clasificación de Baltimore.
6- El Togaviridae es una familia de virus que incluye los siguientes géneros:
Género Alphavirus; especies típicas: Sindbis virus, Virus de la encefalitis equina oriental, Virus de la encefalitis equina occidental, Virus de la encefalitis equina venezolana, Virus del Río Ross, Virus O'nyong'nyong.
Género Rubivirus; especies típicas: Rubéola virus.
7- Los Coronavirus son un género de virus ARN de vertebrados de Coronaviridae.1 Son virus envueltos con un genoma de ARN de cadena sencilla con polaridad positiva y simetría helicoidal. El tamaño genómico de los coronavirus varía entre 16 a 31 kilobases, un virus de extraordinaria longitud.
8- Bunyavirus, género de virus ARN de vertebrados de la familia Bunyaviridae.
9- Papovaviridae es una familia de virus pequeños (apróx. 50 nm) infectivos para una gran variedad de animales, incluyendo los humanos. Su nombre implica las patología que causa: papilomavirus +
Continuacion
ResponderEliminar10- Los paramixovirus son miembros de la familia de virus paramyxoviridae, que poseen un genoma con ARN de cadena sencilla de polaridad negativa como ácido nucleico; albergan dicha información genética en una cápside cubierta de envoltura viral y estructuralmente definida por una simetría helicoidal, de un tamaño de 125 a 250 x 18 nm, con envoltura, pleomórfico, y un filamentoso nucleocápside; y por ensamblar los viriones maduros en el citoplasma como compartimento celular
11- Los Orthomyxoviridae son una familia de virus RNA que infectan a los vertebrados. Incluyen a los virus causantes de la gripe.
12- Un retrovirus es un virus de ARN que se replica en una célula huésped a través del proceso de transcripción inversa . En primer lugar, utiliza su propio transcriptasa inversa enzima para producir ADN de su ARN del genoma, la inversa de la pauta habitual, por lo tanto retro (hacia atrás).
13- Flavivirus : es un género de virus ARN pertenecientes a la familia Flaviviridae. Los Flavivirus son virus con envoltura, la simetría de la nucleocápside icosaedrica, y cuyo material genético reside en una única cadena de ARN de polaridad positiva.
14- Parvovirus, a veces truncada a "parvo", es a la vez el nombre común en Inglés casualmente se aplica a todos los virus de la Parvoviridae familia taxonómica y también el nombre taxonómico del parvovirus género dentro de la Parvoviridae familia.
15- Los adenovirus (Adenoviridae) son una familia de virus que infectan tanto humanos como animales. Son virus no encapsulados de ADN bicatenario que pueden provocar infecciones en las vías respiratorias, conjuntivitis, cistitis hemorrágica y gastroenteritis. Los adenovirus también se utilizan para obtener ADN para la terapia génica.
16- El Arenavirus es un género de virus de la familia Arenaviridae.La especie tipo es el virus de la coriomeningitis linfocítica (LCMV); es también de las spp. responsables de la fiebre Lassa. Arena, viene del latín con la familia de términos del árido.
17- Los herpesvirus o virus de la familia Herpesviridae, deben su nombre al término griego herpein (ἕρπειν), reptar o arrastrar, haciendo alusión a la facultad de estos agentes infecciosos microscópicos de ser fácilmente contagiados y transmitidos de una persona a la otra y de recurrencia crónica. Los herpesvirus forman una familia de virus divididos en tres subfamilias: alpha, beta y gammaherpesvirinae, ubicados con base en la arquitectura del virión y ciertas propiedades biológicas comunes.
18- Poxviridae es una familia de virus de ADN relacionados entre sí llamados poxvirus, infectivos para animales vertebrados (8 de los 11 géneros, por ejemplo, poxvirus de la gallina) y a los invertebrados (3 géneros). Tienen un genoma ADN bicatenario y por lo tanto pertenece al Grupo I de la Clasificación de Baltimore.
19- Filoviridae es una familia de virus del orden Mononegavirales. Se caracterizan por poseer como genoma RNA monocatenario de sentido negativo, y por infectar a primates. Agrupa, entre otros, al virus Ébola y al virus Marburg.
Cristian Rafael Guerrero Mendoza Mat 89275
ResponderEliminarDiagnostico de la Virosis
Los virus son organismos tan pequeños, que su tamaño les permite pasar a través de los filtros que retienen bacterias. Las características básicas para identificar a un virus son: la composición del ácido nucleico, la disposición de la cápside, la presencia o ausencia de una envoltura, el tamaño del virus y el huésped.
MUESTRAS PARA DIAGNOSTICO:
Las muestras deben tomarse precozmente en la fase aguda de la infección. Deben transladarse al laboratorio a la mayor brevedad posible, en medio de transporte. Las muestras más comunes son:
• Torundas nasofaríngeas: adecuados para el diagnóstico de enterovirus, adenovirus, y herpes simplex. Las nasofaringeas se prefieren para el virus respiratorio sincicial y la mayoria de los otros respiratorios. Las nasales son las mejores para los rinovirus.
• Aspirados faríngeos: son mejores que los hisopos (torundas) nasofaringeos, pero estos últimos son mas prácticos.
• Torundas rectales y muestras de heces: Aquí se muestran virus no cultivables como rotavirus, o adenovirus, que se detectan por microscopía electrónica o con técnicas de detección de antígenos.
• Secreciones, Sangre o Líquido cefalorraquídeo.
• En algunas virosis excepcionalmente el diagnóstico solo puede efectuarse en Biopsia de cerebro, como en la encefalitis por herpes simplex; en Huéspedes inmunodeprimidos y con citomegalovirus, el virus solo puede identificarse en el órgano afectado.
DIAGNOSTICO VIRAL
Observación del efecto citopático, en cultivo de tejidos inoculados con ciertos virus. La etiología de un determinado síndrome viral puede demostrarse recuperando el agente causal en tejidos celulares de diversos tipos (por ej: riñón de mono, pulmón humano fetal, amnios humano o células tumorales humanas). Los huevos embrionados actualmente solo se utilizan para cultivar el virus cuando se desea preparar vacunas. El empleo de animales de experimentación es excepcional en el diagnóstico.
Cristian Rafael Guerrero Mendoza Mat 89275
ResponderEliminarReplicación de los virus
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada.
La única característica de la multiplicación viral consiste en que, poco después de su interacción con una célula huésped, el virión infectante se desintegra y pierde la infecciosidad que se puede medir. Esta fase del ciclo celular se llama periodo de eclipse; su duración varía dependiendo del virus y de la célula huésped en particular, y termina con la formación de las primeras partículas virales descendientes infecciosas. El periodo de eclipse es, en realidad, de intensa actividad sintética, puesto que la célula se redirige hacia la satisfacción de las necesidades del "pirata" viral. En algunos casos, tan pronto como el ácido nucleico viral penetra a la célula huésped, el metabolismo celular es recanalizado exclusivamente hacia la síntesis de nuevas partículas virales. En otros casos, los procesos metabólicos de la célula huésped no se alteran significativamente, aunque la célula sintetiza proteínas virales y ácidos nucleicos.
Los virus han desarrollado diversas estrategias para lograr la multiplicación en las células huéspedes parasitadas por ellos. Aunque los detalles varían de un grupo a otro, el patrón general de los ciclos de replicación es semejante.
Fijación, penetración y pérdida de la cubierta:
La primera etapa de la infección viral es la interacción de un virión con un sitio receptor específica de la superficie celular. Las moléculas receptoras difieren para los distintos virus, y son proteínas en algunos casos (por ejemplo, picornavirus) y oligosacáridos en otros (por ejemplo, ortovirus y paramixovirus). La presencia o la ausencia de receptores desempeña una función determinante de primer orden en el tropismo celular y la patogenia viral; por ejemplo, el poliovirus es capaz de fijarse sólo en las células del sistema nervioso central y las vías digestivas de los primates. Se cree que la fijación en receptores refleja homologías configurativas fortuitas entre la estructura de la superficie del virión y un componente de la superficie celular. Por ejemplo, el virus de la inmunodeficiencia humana (HIV) se une al receptor CD4 en las células del sistema inmunitario y se cree que el virus de la rabia interactúa con los receptores de acetilcolina y que el virus de Epstein-Barr reconoce al receptor para el tercer componente del complemento en las células B. Es probable que cada célula susceptible posea por lo menos 100,000 sitios receptores para un virus dado.
Después de la fijación, la partícula viral entra en la célula. Esta etapa se conoce como penetración, viropexia o fagocitosis. En algunos sistemas, se realiza por endocitosis mediada por receptores, con captación de las partículas virales ingeridas dentro de endosomas. Están menos claros los detalles de la penetración en otros sistemas. Ocurre perdida de la cubierta de manera concomitante con la penetración o poco después de la misma. La pérdida de la cubierta, o descubrimiento, es la separación física del ácido nucleico viral (o, en algunos casos, de las nucleocápsides inteARNs) a partir de los componentes estructurales externos del virión. En este momento se pierde la infecciosidad del virus original.
2011-0119 dios te bendiga
ResponderEliminarciclo litico de un bacteriófago.formado por acido nuceico ADN y capsida compleja.fijacion o adsorcion: se fijan inicialmente a traves de las puntas de las fibras cudales mediante enlaces quimicos y posterormente de forma mecanica al clavar las pespinas basales en la pared bacteriana.
penetracion: el bacteriofago mediante enzimas lisozimas situadas en su placa basal perrfora la pared cecular y luego contrae la vaina e introduce el eje tubular de modo que el adn pasa directamente al citoplasma
Continuación
ResponderEliminarLos virus son los únicos agentes infecciosos en los que la disolución del agente infectante es una etapa obligatoria de la vía de la replicación.
Síntesis de los componentes virales:
Poco después de quedar descubierto el genoma viral, se inicia la fase sintética del ciclo de replicación del virus. El aspecto esencial de la replicación viral consiste en que deben transcribirse los ARNm específicos del ácido nucleico viral para que se exprese y duplique con éxito la información genética. Una vez que se logra esta etapa, los virus recurren a los componentes de la célula para traducir el ARNm. Diversas clases de virus emplean vías diferentes para sintetizar los ARNm según la estructura del ácido nucleico viral. Algunos virus (por ejemplo, rhabdovirus, ortomixovirus y paramixovirus) poseen ARN polimerasas que sintetizan a los ARNm. Los virus ARN de este tipo se llaman virus de tira negativa (de sentido negativo), puesto que su genoma de ARN de una sola tira es complementario del ARNm, el cual se designa por acuerdo general como de tira positiva (o de sentido positivo).
Durante la replicación viral se sintetizan todas las macromoléculas especificadas del virus en una secuencia muy organizada. En algunas infecciones por virus, notablemente las producidas por virus de doble tira que contienen ADN, se sintetizan proteínas virales tempranas poco después de la infección y aparecen proteínas tardías sólo durante la fase tardía del proceso infeccioso, una vez que ha ocurrido la síntesis del ADN viral. Pueden activarse o no genes tempranos cuando se elaboran los productos tardíos. En contraste, se expresa al mismo tiempo la mayor parte de la información genética de los virus que contienen ARN o toda ella. Además de estos controles temporales, existen también controles cuantitativos puesto que no se elaboran todas las proteínas virales en las mismas cantidades. Las proteínas específicas del virus pueden regular la extensión de la transcripción del genoma o la traducción del ARNm viral.
Cristian Rafael Guerrero Mendoza Mat 89275
ResponderEliminarAdhesión
La adhesión es la propiedad de la materia por la cual se unen y plasman dos superficies de sustancias iguales o diferentes cuando entran en contacto, y se mantienen juntas por fuerzas intermoleculares.
La adhesión ha jugado un papel muy importante en muchos aspectos de las técnicas de construcción tradicionales. La adhesión del ladrillo con elmortero (cemento) es un ejemplo claro.
La cohesión es distinta de la adhesión. La cohesión es la fuerza de atracción entre partículas adyacentes dentro de un mismo cuerpo, mientras que la adhesión es la interacción entre las superficies de distintos cuerpos.1
Mecanismos de adhesión
Adhesión mecánica
Los materiales adhesivos rellenan los huecos o poros de las superficies manteniendo las superficies unidas por enclavamiento. Existen formas a gran escala de costura, otras veces a media escala como el velcro y algunos adhesivos textiles que funcionan a escalas pequeñas. Es un método similar a la tensión superficial
Adhesión química
Dos materiales pueden formar un compuesto químico al unirse. Las uniones más fuertes se producen entre átomos donde hay permutación (enlace iónico) o se comparten electrones (enlace covalente). Un enlace más débil se produce cuando un átomo de hidrógeno que ya forma parte de una partícula se ve atraída por otra de nitrógeno, oxígeno o flúor, en ese caso hablaríamos de un puente de hidrógeno.
Adhesión dispersiva
En la adhesión dispersiva, dos materiales se mantienen unidos por las fuerzas de van der Waals: la atracción entre dos moléculas, cada una de las cuales tiene regiones de carga positiva y negativa. En este caso, cada molécula tiene una región de mayor carga positiva o negativa que se une a la siguiente de carga contraria. Este efecto puede ser una propiedad permanente o temporal debido al movimiento continuo de los electrones en una región..
Adhesión electrostática
Algunos materiales conductores dejan pasar electrones formando una diferencia de potencial al unirse. Esto da como resultado una estructura similar a un condensador y crea una fuerza electrostática atractiva entre materiales. mi con diversos materiales les pueden ayudar
Cristian Rafael Guerrero Mendoza Mat 89275
ResponderEliminarPenetración
La penetración es el acto de introducir un elemento dentro de otro elemento.
Imagen:
Muestras los diferentes procesos que experimenta la penetración en estos procesos correspondientes a los virus.
Cristian Rafael Guerrero Mendoza Mat 89275
ResponderEliminarPenetración de los Bacteriófagos
Los bacteriófagos (también llamados fagos -del griego φαγητόν (phagētón), «alimento, ingestión») son virus que infectan exclusivamente a lasbacterias.
Al igual que los virus que infectan células eucariotas, los fagos están constituidos por una cubierta proteica o cápside en cuyo interior está contenido su material genético, que puede ser ADN o ARN de simple o doble cadena, circular o lineal (en el 95% de los fagos conocidos es ADN de doble cadena), de 5.000 a 500.000 pares de bases. El tamaño de los fagos oscila entre 20 y 200 nm aproximadamente.
Los fagos son ubicuos y pueden ser encontrados en diversas poblaciones de bacterias, tanto en el suelo como en la flora intestinal de los animales. Uno de los ambientes más poblados por fagos y otros virus es el agua de mar, donde se estima que puede haber en torno a 109partículas virales por mililitro, pudiendo estar infectadas por fagos el 70% de las bacterias marinas.
Historia
La historia del descubrimiento de los bacteriofagos ha sido objeto de muchos debates, los cuales incluyen una controversia sobre quién fue su descubridor. En 1913, el bacteriólogo británicoFrederick Twort descubrió un agente bacteriolítico que infectaba y mataba a las bacterias, pero, no fue capaz de saber qué era exactamente dicho agente, señalando en una de sus hipótesis que, entre otras posibilidades, podría tratarse de un virus. No fue sino hasta 1917, cuando el microbiólogo canadiense Félix d'Herelle anunció el descubrimiento de "un invisible antagonista microbiano del bacilo de la disentería", al cual afirmó que se trataba de un virus al cual llamó bacteriófago. Los trabajos de d'Herelle iniciaron desde 1910 y en contraste con Twort y algunos otros científicos que habían reportado fenómenos similares, tuvo pocas dudas sobre la naturaleza del fenómeno que estaba observando y afirmó que se trataba de un virus que parasitaba a la bacteria. El nombre bacteriófago lo formó de la palabra "bacteria" y "phagein" (comer o devorar, en griego), implicando que los fagos "comen" o "devoran" a las bacterias.
continuacion partes 7 2011-0119 dios te bendiga
ResponderEliminarDiagnostico de la Virosis
Los virus son organismos tan pequeños, que su tamaño les permite pasar a través de los filtros que retienen bacterias. Las características básicas para identificar a un virus son: la composición del ácido nucleico, la disposición de la cápside, la presencia o ausencia de una envoltura, el tamaño del virus y el huésped.
MUESTRAS PARA DIAGNOSTICO:
Las muestras deben tomarse precozmente en la fase aguda de la infección. Deben transladarse al laboratorio a la mayor brevedad posible, en medio de transporte. Las muestras más comunes son:
• Torundas nasofaríngeas: adecuados para el diagnóstico de enterovirus, adenovirus, y herpes simplex. Las nasofaringeas se prefieren para el virus respiratorio sincicial y la mayoria de los otros respiratorios. Las nasales son las mejores para los rinovirus.
• Aspirados faríngeos: son mejores que los hisopos (torundas) nasofaringeos, pero estos últimos son mas prácticos.
• Torundas rectales y muestras de heces: Aquí se muestran virus no cultivables como rotavirus, o adenovirus, que se detectan por microscopía electrónica o con técnicas de detección de antígenos.
• Secreciones, Sangre o Líquido cefalorraquídeo.
Mat:2011-0289 Maduracion y salida de virus envueltos y desnudos.
ResponderEliminarLos virus cubiertos maduran por un proceso de gemación. Se insertan glucoproteínas de cubierta específica del virus en las membranas celulares y, a continuación, las nucleocápsides hacen gemación a través de la membrana a nivel de estos sitios modificados y, al hacerlo así, adquieren su cubierta. La gemación ocurre a menudo a nivel de la membrana plasmática y puede abarcar a otras membranas de la célula. Los virus cubiertos no son infecciosos hasta que han adquirido sus cubiertas. Por lo tanto, no se acumulan de manera típica viriones descendientes infecciosos en el interior de la célula infectada.
La maduración viral es a veces un proceso ineficaz. Pueden acumularse cantidades excesivas de componentes virales y participar en la formación de cuerpos de inclusión en la célula (ver antes). Como resultado de los profundos efectos dañinos de la replicación viral, se desarrollan por último efectos citopáticos celulares y la célula muere. Sin embargo, hay casos en que el virus no lesiona a la célula y se producen infecciones persistentes y prolongadas.os genomas virales recién sintetizados y los polipéptidos de la cápside se ensamblan para formar los virus descendientes. Las cápsides icosaédricas pueden condensarse en ausencia de ácido nucleico, en tanto que las nucleocápsides de los virus de simetría helicoidal no pueden formarse sin ARN viral.
No existen mecanismos especiales para la liberación de virus carentes de cubierta; las células infectadas acaban por experimentar lisis y liberar las partículas virales.
Adhesión
ResponderEliminarLa adhesión es la propiedad de la materia por la cual se unen y plasman dos superficies de sustancias iguales o diferentes cuando entran en contacto, y se mantienen juntas por fuerzas intermoleculares.
Mecanismos de adhesion
La cohesión es la causa de que el agua forme gotas, la tensión superficial hace que se mantengan esférica y la adhesión la mantiene en su sitio.
Las gotas de agua son más planas sobre la flor de Hibiscus ya que tienen mejor adhesión.
Cinco mecanismos han sido propuestos para explicar por qué un material se adhiere a otro.
Adhesión mecánica
Los materiales adhesivos rellenan los huecos o poros de las superficies manteniendo las superficies unidas por enclavamiento. Existen formas a gran escala de costura, otras veces a media escala como el velcro y algunos adhesivos textiles que funcionan a escalas pequeñas. Es un método similar a la tensión superficial.
Adhesión química
Dos materiales pueden formar un compuesto químico al unirse. Las uniones más fuertes se producen entre átomos donde hay permutación (enlace iónico) o se comparten electrones (enlace covalente). Un enlace más débil se produce cuando un átomo de hidrógeno que ya forma parte de una partícula se ve atraída por otra de nitrógeno, oxígeno o flúor.
Penetración de los Bacteriófagos
Los virus son moléculas de DNA o RNA rodeadas por una envoltura proteica que necesitan células viables para poder replicarse. Los virus utilizan la maquinaria metabólica de las células para sintetizar su material genético y proteínas de la envoltura. Existen distintos tipos de virus que pueden infectar células procariontes o células eucariontes. Los bacteriófagos o fagos son virus que se reproducen en células procariontes.
Replicacion:
Los virus sólo se multiplican en células vivientes. La célula huésped debe proporcionar la energía y la maquinaria de síntesis, también los precursores de bajo peso molecular para la síntesis de las proteínas virales y de los ácidos nucleicos. El ácido nucleico viral transporta la especificidad genética para cifrar todas las macromoléculas específicas virales en una forma altamente organizada
Mat:2011-0289 SALIDA DE UN VIRUS ENVUELTO.
ResponderEliminarLos virus pueden ser liberados mediante lisis celular, o, si son envueltos, pueden yemar de las células. Los virus que yeman no necesariamente destruyen la célula. Por tanto, algunos de estos virus son capaces de instaurar infecciones persistentes.
No todas las partículas víricas liberadas son infecciosas.
La proporción partículas infecciosas:partículas no-infecciosas varía según el virus y las condiciones de crecimiento.
Los virus envueltos (como el VIH) son liberados de la célula huésped por gemación. Durante este proceso, el virus adquiere su envoltura, que es una parte modificada de la membrana plasmática del huésped.
Maduración: de produce el ensamble de los ácidos nucleicos neoformados con las proteínas de la cápside.
Liberación: los nuevos viriones salen de la célula por lisis de la misma, como el virus polio; o bien por brotación a través de la membrana celular, como los paramixovirus o arenavirus; o de la membrana nuclear, como los herpesvirus o del retículo endoplásmico, como los togavirus.
continuacion 2011-0119 dios te bendiga
ResponderEliminarMaduración y ensamblaje
Cuando se han sintetizado un número crítico de varios componentes virales, éstos se ensamblan en partículas víricas maduras en el núcleo y/o citoplasma de la célula infectada. En este ensamblaje y al contrario de lo que ocurre en los bacteriófagos, no están implicados enzimas biosintéticos especiales, sino que ocurre de forma espontánea debido a la gran afinidad de la cápsida con el ácido nucleico viral. La maduración se puede definir como la fase de la infección viral durante la cual los componentes estructurales del virión se producen y ensamblan con el nuevo ácido nucleico viral para formar la nucleocápsida.
Salida de un Virus Envuelto
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula
Mat:2011-0289 =RETROVIRUS
ResponderEliminarFármacos Antivirales:
Las drogas antirretrovirales actúan sobre diferentes pasos necesarios para la replicación viral; hasta la fecha, las más extendidas son aquellas drogas que actúan sobre la transcriptasa inversa y las drogas que actúan sobre las proteasas; aunque ya existen fármacos capaces de inhibir otros sitios tales como los inhibidores de la fusión y los de la integrasa.
Los Inhibidores de la Reversotranscriptasa (IRT) alteran la función de la enzima llamada reversotranscriptasa que es la que utiliza el VIH para cambiar su mensaje genético a una forma que pueda ser fácilmente insertada dentro del núcleo de la célula infectada. Es la encargada de hacer una copia en ADN del ARN viral de manera que éste pueda insertarse en el ADN celular, mediante la acción de las integrasas, y así producir proteínas del VIH necesarias para la auto reproducción viral. Los IRT nucleósidos y los nucleótidos necesitan ser fosforilados para luego interactuar con un sustrato en el sitio de unión a la enzima y, cuando es incorporado, entonces termina la elongación de la cadena de ADN; mientras que los IRT no nucleósidos no necesitan ninguna conversión metabólica y bloquean directamente la reacción con la reverso transcriptasa por una interacción específica con el sitio de unión “sin sustrato” de la reversotranscriptasa del VIH.
Los Inhibidores de las Proteasas (IP) son fármacos con una potente actividad antiviral. La proteasa del VIH es requerida para el clivaje del polipéptido gag-pol en sus partes funcionales y la inhibición de este clivaje trae como consecuencia una disminución en la producción de viriones maduros. O sea, estos medicamentos se parecen a los trozos de la cadena de proteína que la proteasa corta normalmente. Al adherirse a la proteasa, los IP previenen que la proteasa corte las cadenas largas de proteínas y enzimas en trozos más cortos que el virus necesita para reproducirse en nuevas copias de sí mismo. Aunque las cadenas largas no hayan sido cortadas en los trocitos correctos, las nuevas copias del VIH aún se construyen y continúan empujando a través de la membrana de la célula infectada. Pero estas nuevas copias del VIH son “defectuosas”, ya que no han sido formadas completamente, por lo tanto no pueden continuar infectando otras células.(1,2,3,4)
INHIBIDORES DE FUSION
Los inhibidores de la entrada evitan el ingreso del VIH a las células sanas, se adhieren a las proteínas de superficie de las células T o a las proteínas de superficie del VIH. Para que el VIH es una y pueda entrar a las células T, las proteínas de la capa externa del VIH deben unirse a las proteínas de superficie de las células T.
Los inhibidores de la entrada evitan que este proceso ocurra. Algunos inhibidores de la entrada se adhieren a las proteínas gp120 ó gp41 de la capa externa del VIH. Otros inhibidores utilizan la proteína CD4 ó los receptores CCR5 ó CXCR4 de la superficie de las células T. Si los inhibidores de la entrada logran adherirse eficientemente a las proteínas, éstas quedan bloqueadas y entonces el VIH no puede unirse a la superficie de las células T e ingresar a las mismas.
MECANISMO DE ACCION DE LOS RETROVIRALES
Los antivirales son un tipo de medicamento usado para el tratamiento de infecciones producidas por virus. Tal como los antibióticos (específicos para bacteria), existen antivirales específicos para distintos tipos de virus. No sin excepciones, son relativamente inocuos para el huésped, por lo que su aplicación es relativamente segura. Deben distinguirse de los viricidas, que son compuestos químicos que destruyen las partículas virales presentes en el medio ambiente.
Su eficacia (y a veces su toxicidad) están relacionadas con la competencia inmunológica del paciente.
Su actividad es restringida al actuar sobre un “órgano diana” muy específico.
Desarrollo de Resistencias.
La mayoría solo son eficaces durante la fase de Replicación viral y no sobre los virus Latentes.
La utilización óptima de los antivirales precisa de la realización de un diagnóstico clínico correcto.
Mat:2011-0289 =BIONES ACELULARES
ResponderEliminarEl término acelular significa literalmente "sin células". Se aplica principalmente a los virus, pero también a provirus y viroides, que son entidades biológicas no formadas por células, a diferencia de bacterias, compuestas por células procariotas, y de hongos, vegetales y animales, formados por células eucariotas.
LOS VIROIDES I y II
son agentes infecciosos que, al igual que los virus, tienen un ciclo extracelular que se caracteriza por la inactividad metabólica y unciclo intracelular en el que causan infección al huésped susceptible, pero que a diferencia de los virus, los viroides no poseen proteínas ni lípidos y están constituidos por una cadena cíclica corta de ARN, circular o con forma de varilla, (que no codifica proteínas).1 Es importante decir que tanto su forma intracelular como extracelular son las mismas (ARN desnudo), los mecanismos por los cuales éstos logran causar infección están relacionados con la autocatálisis de su material genético.
Estas pequeñas moléculas de ARN circular de cadena sencilla formadas por unos pocos cientos de nucleótidos. Carecen de cápsida y su tamaño es una milésima parte del de los virus más pequeños.
- Aparecen con mayor abundancia en el núcleo de las células infectadas.
- Sólo han sido detectados en plantas, en las que producen una gran variedad de enfermedades y pueden ser transmitidos tanto horizontal como verticalmente.
- No existen ninguna evidencia de que los viroides son traducidos a proteínas, ni se conoce cómo causan las enfermedades. Lo que sí se sabe es que son replicados por los enzimas del huésped.
- Algunas semejanzas entre la secuencia de nucleótidos de los viroides y ciertos secuencias del genoma de las plantas
no traducidas (intrones) sugieren que los viroides evolucionaron a partir de los intrones.
2011-0119 dios te bendiga contuacion
ResponderEliminarMaduración y Salida de Virus Desnudos y Envueltos
Virus envueltos
(A) Entran por fusión con la membrana plasmática. Algunos virus envueltos se fusionan directamente con la membrana plasmática. Por lo que los componentes internos del virión son inmediatamente llevados al citoplasma celular. (Figura 1).
(B) Entada vía endosomas en la superficie celular (figura 2)
Algunos virus envueltos requieren de un pH ácido para que ocurra la fusión y no son capaces de fusionarse directamente con la membrana celular. Estos virus acogidos o tomados mediante invaginación de la membrana en endosomas. A medida que los endosomas se acidifican la actividad de fusión, hasta ahora latente, de las proteínas virales se activa, y la membrana del virión se fusiona con la membrana del endosoma. Esto resulta en el transporte de los componentes internos del virus hacia el citoplasma de la célula.
Los antivirales son un tipo de medicamento usado para el tratamiento de infecciones producidas por virus. Tal como los antibióticos (específicos para bacteria), existen antivirales específicos para distintos tipos de virus. No sin excepciones, son relativamente inocuos para el huésped, por lo que su aplicación es relativamente segura. Deben distinguirse de los viricidas, que son compuestos químicos que destruyen las partículas virales presentes en el medio ambiente
Mat:2011-0289 =Otros biones Acelulares
ResponderEliminarUn virus satélite es un agente subvírico formado por ácido nucleico y que depende de la coinfección de la célula huésped por un virus (helper o máster) para reproducirse; obteniendo de ellos lasenzimas faltantes en su genoma, las cuales son necesarias para lograr replicarse.
Un virusoide o ARN satélite es una pequeña molécula patógena de ARN (ácido ribonucleico) que se encuentra encapsulada en ciertos virus de plantas con secuencias de ARN de mayor tamaño, lo que se denomina un virus asistente, el cual hospeda al virusoide y es necesario para que éste produzca una infección exitosa.
La McIntosh Roja o Mac es un tipo de manzana de piel roja y verde. Se recoge a finales de septiembre y es el cultivo más popular de Nueva Inglaterra, muy conocida por la compota rosada que se hace a partir de manzanas McIntosh sin pelar.
Cada manzana de McIntosh tiene un linaje directo a un solo árbol descubierto en 1796 por John McIntosh en su granja en Dundela, una aldea situada en el condado de Dundas en la provincia canadiense de Ontario, cerca de Prescott.
Los plásmidos son fragmentos extracromosómicos de ácidos nucléicos (ADN o ARN) que aparecen en el citoplasma de algunos procariotas. Son de tamaño variable aunque menor que el cromosoma principal.
Mat:2011-0289 =Priones I
ResponderEliminarLos priones son los agentes causantes de un grupo de patologías neurodegenerativas letales características de mamíferos, también conocidas como encefalopatías espongiformes transmisibles. Estos agentes son capaces de propagarse dentro de un mismo huesped causando una lesión espongiótica y de transmitirse de huesped a huesped con elevados tiempos de incubación. A diferencia de virus y viroides, son resistentes a tratamientos inactivantes de ácidos nucléicos, pero comparten con éstos la existencia de una variabilidad de inóculos dentro de la misma especie (diferenciables por el patrón de la lesión y la magnitud del tiempo de incubación) y de una infectividad sujeta a barrera de especie (Chandler, 1961; Alper y cols., 1967; Hunter, 1972; Prusiner, 1982; Bruce y Fraser, 1991; Bessen y Marsh, 1992).
La búsqueda de la entidad molecular constitutiva de este agente reveló como componente mayoritario, si no único, una proteína: PrPSc, proteína del prion de scrapie),y la ausencia de un ácido nucleico específico (Prusiner 1982, 1991). Con estas premisas estructurales unidas a la capacidad de infección, Prusiner acuña el término prion (particula infecciosa de naturaleza proteica) para diferenciarlo de virus y viroides. Dadas las características poco convencionales de los priones se han elaborado numerosas hipótesis sobre su estructura (Dickinson y Outram, 1988; Prusiner, 1991; Weissmann, 1991). En la actualidad la hipótesis con mayor grado de aceptación es la conocida como "sólo proteína", delineada inicialmente por Griffith (1967) y, formalmente enunciada y actualizada por Prusiner (1991, 1997). Tras la descripción de proteínas de diferente secuencia pero similar comportamiento en levadura así como los avances interdisciplinares realizados en el conocimento de estos agentes en la última década, el concepto de prion ha sido acotado adquiriendo una defición más precisa y generalizable (Wicker y Masison, 1996; Lindquist S., 1997; Prusiner, 1997). Así, se denomina prion a la forma alterada de una proteína celular funcional (PrP en mamíferos) que ha podido perder su función normal pero que ha adquirido la capacidad de transformar la forma normal en patológica.
Mat:2011-0289 =Priones II
ResponderEliminarHan sido definidos como pequeñas partículas proteináceas que son resistentes a la inactivación por procedimientos que modifican los ácidos nucléicos.
Otra definición de prion es la forma alterada de una proteína celular funcional que ha podido perder su función normal pero que ha adquirido la capacidad de transformar la forma normal en patológica.
Los mamíferos tienen en su genoma genes que codifican la “proteína priónica”(PrP), y esta proteína en condiciones normales tiene una configuración espacial de tipo hélices alfa, que es susceptible a la lisis por proteasas y no es patógena. Los priones no son seres vivos ni tienen la capacidad de infección como tal.
La patogénesis de estas proteínas se produce cuando tienen bien una mutación (esporádica o adquirida genéticamente) o bien un cambio conformacional inducido por PrP-Sc (patógena) de otro individuo.
El cambio conformacional o las mutaciones hacen que la PrP tenga una estructura tridimensional diferente constituida por hélices beta con agregados amorfos insolubles (PrP-Sc), de manera que se hace imposible su lisis por las proteasas, produciéndose su acúmulo en los lisosomas de las células de los tejidos afectados y originando así las enfermedades priónicas.
Juan Miguel Mazara Simon
ResponderEliminar2011-0444
Martes, 3:00-5:00 PM
Virología es el estudio de virus y virus como agentes: clasificación y evolución, su estructura, sus formas de infectar y explotar las células para la reproducción del virus, las enfermedades que causan, las técnicas para aislar y la cultura de ellos y su uso en la investigación y terapia.
Virología es considerada una parte de la microbiología o de patología.
Una importante rama de la Virología es la clasificación de virus. Virus pueden clasificarse según la célula huésped que infectan: virus animales, virus de plantas, hongos virus y bacteriófagos (virus que infectan bacterias, que son los virus más complejos).
Otra clasificación utiliza la forma geométrica de su cápside (a menudo una hélice o un icosaedro) o la estructura del virus (por ejemplo, la presencia o la ausencia de un envolvente de lípidos). Gama de virus en tamaño de unos 30 nm a aproximadamente 450 nm, lo que significa que la mayoría de ellos no puede verse con microscopios de luces.
La forma y la estructura del virus ha sido estudiado por cristalografía de rayos X, microscopía electrónica y RMN.
El sistema de clasificación más útil y más ampliamente utilizado distingue virus segun el tipo de ácido nucleico que utilizan como material genético y el método de replicación viral que emplean coaxial células huésped en la producción de más virus:
Virus de ADN (divididas en virus de ADN bicatenario y los virus de ADN monocatenario mucho menos común),
Virus ARN (dividieron en virus de ARN monocatenario de sentido positivo, virus de ARN monocatenario de sentido negativo y los virus de ARN bicatenario mucho menos común),
invertir transcribir virus (virus de ADN de transcripción inversa de bicatenario y monocatenario transcripción inversa RNA virus incluyendo retrovirus).
Además virólogos también estudian '' partículas subvirales '', incluso más pequeñas que los virus infecciosas entidades: viroides (desnudas circulares moléculas de ARN infectando plantas), satélites (moléculas de ácido nucleico con o sin una cápside que requieren un virus auxiliar para la infección y reproducción) y priones (proteínas que pueden existir en una conformación patológica que induce a otras moléculas del prión para asumir esa misma conformación).
El último informe del Comité Internacional de taxonomía de virus (2005) muestra 5450 virus, organizados en más de 2.000 especies, 287 géneros y 73 familias 3 pedidos.
Los taxones en Virología no son necesariamente monofiléticos. De hecho, las relaciones evolutivas de los distintos grupos de virus no están claras, y existen tres hipótesis sobre su origen:
Virus surgieron de materia no viviente, por separado y en paralelo a otras formas de vida, posiblemente en forma de autorreplicación RNA ribozimas similares a viroides.
Virus surgieron de la anteriores, más competentes celulares formas de vida que se convirtieron en parásitos a las células del huésped y posteriormente perdió la mayoría de sus funciones; ejemplos de tales diminutos parásitos procariotas son Mycoplasma y Nanoarchaeum.
Virus surgieron como partes del genoma de las células, probablemente transposones o plásmidos, que adquirieron la capacidad para "romper libre" desde el host de celda y infectan otras células.
EliminarPor supuesto es posible que diferentes alternativas se aplican a grupos de virus diferente.
De particular interés aquí es mimivirus, un virus gigante que infecta a las amebas y lleva gran parte de la maquinaria molecular tradicionalmente asociada con las bacterias. ¿Es una versión simplificada de una Célula procariota parasitaria o se originó como un simple virus que adquirido genes de su host?
La evolución del virus, que a menudo se produce en concierto con la evolución de sus anfitriones, se estudia en el campo de la evolución viral.
Mientras que el virus se reproducen y evolucionan, ellos no participan en el metabolismo y dependen de una célula huésped para la reproducción. La cuestión debatida a menudo de si están vivos o no es una cuestión de definición que no afectan la realidad biológica de los virus.
Un virus (de la palabra latina virus, toxina o veneno) es una entidad biológica capaz de autorreplicarse utilizando la maquinaria celular, es un agente potencialmente patógeno compuesto por una cápside (o cápsida) de proteínas que envuelve al ácido nucléico, que puede ser ADN o ARN. Esta estructura puede a su vez estar rodeada por la envoltura vírica, una capa lipídica con diferentes proteínas, dependiendo del virus.
El estudio de los virus ha sido muy complejo para los científicos, ya que ellos tienen la capacidad de cambiar permanentemente y esto dificulta su control y eliminación. Producen enfermedades como sarampión, gripe, viruela, poliomielitis y un mal que afecta a muchas personas en el mundo: el síndrome de inmuno deficiencia adquirida o sida.
El ciclo vital de un virus siempre necesita de la maquinaria metabólica de una célula para poder replicar su material genético, produciendo muchas copias del virus original. En dicho proceso reside la capacidad destructora de los virus, ya que pueden perjudicar a la célula hasta destruirla. Pueden infectar células eucarióticas o procarióticas (en cuyo caso se les llama bacteriófagos, o simplemente fagos). Algunos indicios parecen demostrar que existen virus que infectan a otros virus (llamados viroides).
Se puede agrupar las características definitorias de los virus en torno a tres cuestiones: su tamaño, el hecho de que sean cristalizables y el hecho de que sean parásitos intracelulares o microcelulares obligados. Estas tres cuestiones colocan a los virus en la frontera entre lo vivo y lo inerte.
Tamaño de los virus
Los virus son estructuras extraordinariamente pequeñas. Su tamaño oscila entre los 24 nanómetros del virus de la fiebre aftosa a los 300 nanómetros de los poxvirus.
Su pequeño tamaño explica lo tardío del descubrimiento de estos agentes. La primera referencia sobre la existencia de los virus se debe al botánico ruso Dimitri Ivanovski en 1892. Este investigador buscaba el agente causante de la enfermedad denominada mosaico del tabaco, y llegó a la conclusión de que debía tratarse de una toxina o de un organismo más pequeño que las bacterias, pues el agente atravesaba los filtros que retenían las bacterias. Denominó a estos agentes patógenos virus filtrables.
EliminarEn 1897, el microbiólogo holandés Martinus Beijerink realizó experimentos similares a los de Ivanovski, y llegó a desechar la idea de las toxinas, pues se trataba de un agente capaz de reproducirse, ya que mantenía su poder infeccioso de unas plantas a otras, sin diluirse su poder patógeno. Poco después, los microbiólogos alemanes Frederick Loeffler y Paul Frosch descubrieron que la fiebre aftosa del ganado era producida por un virus filtrable que actuaba como un agente infeccioso.
En la década de los 30, con el uso de filtros de tamaño de poro inferior, con las técnicas de cultivo celular in vitro que permitían la obtención de gran cantidad de virus, con la ultracentrifugación y finalmente con el microscopio electrónico y la difracción de rayos X, se logró visualizar a estos agentes.
Cristalización
Los virus son cristalizables, como demostró W. Stanley en 1935. Esto significa que las partículas víricas tienen formas geométricas y que son idénticas entre sí, lo cual las separa de la irregularidad característica de los organismos, y las acerca a las características de los minerales.
Parásitos intracelulares obligados
Los virus son parásitos intracelulares obligados. Desde los años treinta se sabe que los virus se componen principalmente de ácido nucleico y proteínas, estas últimas forman la cápside, que se conoce también como envoltura proteínica. Esto quiere decir que necesitan un huésped, ya que en vida libre no sobreviven. Se sabe que los virus pueden vivir alrededor de unos cuarenta días sin que tengan algún huésped en que se continúen reproduciendo.
También se han encontrado virus que presentan lípidos, aunque estos son tomados de la célula que infectan. Hasta ahora todos los virus que se conocen presentan un solo tipo de ácido nucleico (ya sea ADN ó ARN), el cual puede ser de una ó de dos cadenas y puede ser segmentado. Para que el ácido nucleico pueda replicarse, necesita utilizar la maquinaria encimática y estructural de una célula viva, y por otra parte, solamente dentro de una célula viva tienen los virus las funciones de autoconservación, que junto con la reproducción, caracterizan a los seres vivos. Esta condición es la causa de que muchísimos virus sean conocidos como gérmenes patógenos que producen enfermedades en plantas y animales, e incluso en las bacterias.
Estructura de los virus
Un virus está compuesto de una molécula de ácido nucleico y una envoltura proteínica. Ésta es la estructura básica de un virus, aunque algunos de ellos pueden añadir a esto la presencia de alguna enzima, bien junto al ácido nucleico, como la transcriptasa inversa de los retrovirus, bien en la envoltura, para facilitar la apertura de una brecha en la membrana de la célula hospedadora.